Репарация
(от лат. reparatio — восстановление) — особая
функция клеток, заключающаяся в
способности исправлять химические
повреждения и разрывы в молекулах ДНК,
повреждённой при нормальном биосинтезе
ДНК в клетке или в результате воздействия
физических или химических агентов.
Осуществляется специальными ферментными
системами клетки. Ряд наследственных
болезней (напр., пигментная ксеродерма)
связан с нарушениями систем
репарации.
(Процесс,
позволяющий живым организмам
восстанавливать повреждения, возникающие
в ДНК, называют репарацией – кратко).
Все
репарационные механизмы основаны на
том, что ДНК — двухцепочечная молекула,
т.е. в клетке есть 2 копии генетической
информации. Если нуклеотидная
последовательность одной из двух цепей
оказывается повреждённой (изменённой),
информацию можно восстановить, так как
вторая (комплементарная) цепь
сохранена.
Процесс
репарации происходит в несколько этапов.
На первом этапе выявляется нарушение
комплементарности цепей ДНК. В ходе
второго этапа некомплементарный
нуклеотид или только основание
устраняется, на третьем и четвёртом
этапах идёт восстановление целостности
цепи по принципу комплементарности.
Однако в зависимости от типа повреждения
количество этапов и ферментов, участвующих
в его устранении, может быть разным.
Очень
редко происходят повреждения, затрагивающие
обе цепи ДНК, т.е. нарушения структуры
нуклеотидов комплементарной пары. Такие
повреждения в половых клетках не
репарируются, так как для осуществления
сложной репарации с участием гомологичной
рекомбинации требуется наличие
диплоидного набора хромосом.
Виды:
1)Прямая
репарация
2)Эксцизионная
репарация
3)Пострепликативная
репарация
Прямая
репарация
— наиболее простой путь устранения
повреждений в ДНК, в котором обычно
задействованы специфические ферменты,
способные быстро (как правило, в одну
стадию) устранять соответствующее
повреждение, восстанавливая исходную
структуру нуклеотидов. Так действует,
например, O6-метилгуанин-ДНК-метилтрансфераза,
которая снимает метильную группу с
азотистого основания на один из
собственных остатков цистеина.
Эксцизионная
репарация (англ. excision — вырезание)
включает удаление повреждённых азотистых
оснований из ДНК и последующее
восстановление нормальной структуры
молекулы. Эксцизионная репарация
(excision repair): процесс с участием ферментативной
системы, которая удаляет короткую
однонитевую последовательность
двунитевой ДНК , содержащей ошибочно
спаренные или поврежденные основания
, и замещает их путем синтеза
последовательности, комплементарной
оставшейся нити.
Эксцизионная
репарация является наиболее распространенным
способом репарации модифицированных
оснований ДНК. Этот тип репарации
базируется на распознавании
модифицированного основания различными
гликозилазами, расщепляющими N-гликозидную
связь этого основания с сахарофосфатным
остовом молекулы ДНК. При этом существуют
гликозилазы, специфически распознающие
присутствие в ДНК определенных
модифицированных оснований
(оксиметилурацила, гипоксантина,
5-метилурацила, 3-метиладенина,
7-метилгуанина и т.д.). Для многих гликозилаз
к настоящему времени описан полиморфизм,
связанный с заменой одного из нуклеотидов
в кодирующей последовательности гена.
Для ряда изоформ этих ферментов была
установлена ассоциация с повышенным
риском возникновения онкологических
заболеваний [ Chen, 2003 ].
Пострепликативная
репарация
Tип
репарации, имеющей место в тех случаях,
когда процесс эксцизионной репарации
недостаточен для полного исправления
повреждения: после репликации с
образованием ДНК, содержащей поврежденные
участки, образуются одноцепочечные
бреши, заполняемые в процессе гомологичной
рекомбинации при помощи белка
RecA.[1]
Пострепликативная
репарация была открыта в клетках E.Coli,
не способных выщеплять тиминовые димеры.
Это единственный тип репарации, не
имеющий этапа узнавания повреждения.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
У этого термина существуют и другие значения, см. Репарация.

Репарация (от лат. reparatio — восстановление) — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённых при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических реагентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней (напр., пигментная ксеродерма) связан с нарушениями систем репарации.
История открытия
Начало изучению репарации было положено работами Альберта Кельнера (США), который в 1948 году обнаружил явление фотореактивации — уменьшение повреждения биологических объектов, вызываемого ультрафиолетовыми (УФ) лучами, при последующем воздействии ярким видимым светом (световая репарация).
Р. Сетлоу, К. Руперт (США) и другие вскоре установили, что фотореактивация — фотохимический процесс, протекающий с участием специального фермента и приводящий к расщеплению димеров тимина, образовавшихся в ДНК при поглощении УФ-кванта.
Позднее при изучении генетического контроля чувствительности бактерий к УФ-свету и ионизирующим излучениям была обнаружена темновая репарация — свойство клеток ликвидировать повреждения в ДНК без участия видимого света. Механизм темновой репарации облучённых УФ-светом бактериальных клеток был предсказан А. П. Говард-Фландерсом и экспериментально подтверждён в 1964 году Ф. Ханавальтом и Д. Петиджоном (США). Было показано, что у бактерий после облучения происходит вырезание повреждённых участков ДНК с изменёнными нуклеотидами и ресинтез ДНК в образовавшихся пробелах.
Системы репарации существуют не только у микроорганизмов, но также в клетках животных и человека, у которых они изучаются на культурах тканей. Известен наследственный недуг человека — пигментная ксеродерма, при котором нарушена репарация.
Томас Линдаль, Азиз Шанкар и Пол Модрич получили Нобелевскую премию по химии 2015 года за исследования в области изучения методов репарации ДНК[1][2].
Источники повреждения ДНК
- Ультрафиолетовое излучение
- Радиация
- Химические вещества
- Ошибки репликации ДНК
- Апуринизация — отщепление азотистых оснований от сахарофосфатного остова
- Дезаминирование — отщепление аминогруппы от азотистого основания
Основные типы повреждения ДНК
- Повреждение одиночных нуклеотидов
- Повреждение пары нуклеотидов
- Двухцепочечные и одноцепочечные разрывы цепи ДНК
- Образование поперечных сшивок между основаниями одной цепи или разных цепей ДНК
- Образование тиминовых димеров
- Кросслинкинг

Устройство системы репарации
Каждая из систем репарации включает следующие компоненты:
- ДНК-хеликаза — фермент, «узнающий» химически изменённые участки в цепи и осуществляющий разрыв цепи вблизи от повреждения;
- ДНКаза (дезоксирибонуклеаза) — фермент, «разрезающий» 1 цепочку ДНК (последовательность нуклеотидов) по фосфодиэфирной связи и удаляющий повреждённый участок: экзонуклеаза работает на концевые нуклеотиды 3` или 5`, эндонуклеаза — на нуклеотиды, отличные от концевых;
- ДНК-полимераза — фермент, синтезирующий соответствующий участок цепи ДНК взамен удалённого;
- ДНК-лигаза — фермент, замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность.
Типы репарации
У бактерий имеются по крайней мере 3 ферментные системы, ведущие репарацию — прямая, эксцизионная и пострепликативная. У эукариот и бактерий также есть особые виды репарации Mismatch [3] и SOS-репарация (несмотря на название данный вид репарации немного различается у бактерий и эукариот [4].
Прямая репарация
Прямая репарация — наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов. Так действует, например, O6-метилгуанин-ДНК-метилтрансфераза, которая снимает метильную группу с азотистого основания на один из собственных остатков цистеина.
Эксцизионная репарация
Эксцизионная репарация (англ. excision — вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы по комплементарной цепи. Ферментативная система удаляет короткую однонитевую последовательность двунитевой ДНК, содержащей ошибочно спаренные или поврежденные основания, и замещает их путём синтеза последовательности, комплементарной оставшейся нити.
Эксцизионная репарация является наиболее распространённым способом репарации модифицированных оснований ДНК. Она базируется на распознавании модифицированного основания различными гликозилазами, расщепляющими N-гликозидную связь этого основания с сахарофосфатным остовом молекулы ДНК. При этом существуют гликозилазы, специфически распознающие присутствие в ДНК определенных модифицированных оснований (оксиметилурацила, гипоксантина, 5-метилурацила, 3-метиладенина, 7-метилгуанина и т. д.). Для многих гликозилаз к настоящему времени описан полиморфизм, связанный с заменой одного из нуклеотидов в кодирующей последовательности гена. Для ряда изоформ этих ферментов была установлена ассоциация с повышенным риском возникновения онкологических заболеваний [ Chen, 2003 ].
Другой тип эксцизионной репарации — эксцизионная репарация нуклеотидов, предназначенная для более крупных повреждений, таких как образование пиримидиновых димеров.
Пострепликативная репарация
Tип репарации, имеющей место в тех случаях, когда процесс эксцизионной репарации недостаточен для полного исправления повреждения: после репликации с образованием ДНК, содержащей повреждённые участки, образуются одноцепочечные бреши, заполняемые в процессе гомологичной рекомбинации при помощи белка RecA[5].
Пострепликативная репарация была открыта в клетках E. coli, не способных выщеплять тиминовые димеры. Это единственный тип репарации, не имеющий этапа узнавания повреждения.
Интересные факты
- Полагают, что от 80 % до 90 % всех раковых заболеваний связаны с отсутствием репарации ДНК[6].
- Повреждение ДНК под воздействием факторов окружающей среды, а также нормальных метаболических процессов, происходящих в клетке, происходит с частотой от нескольких сотен до 1000 случаев в каждой клетке, каждый час[7].
- По сути ошибки в репарации происходят так же часто как и в репликации, а при некоторых условиях даже чаще.
- В половых клетках сложная репарация, связанная с гомологичной рекомбинацией не происходит из-за гаплоидности генома этих клеток[8].
Примечания
- ↑ Нобелевская премия по химии присуждена за починку ДНК // Lenta.Ru. Дата обращения: 7 октября 2015. Архивировано 7 октября 2015 года.
- ↑ Кирилл Стасевич. Как клетка чинит свою ДНК // Наука и жизнь. — 2015. — № 11. — С. 30—38.
- ↑ Kenji Fukui. DNA mismatch repair in eukaryotes and bacteria // Journal of Nucleic Acids. — 2010-07-27. — Т. 2010. — С. 260512. — ISSN 2090-021X. — doi:10.4061/2010/260512. Архивировано 24 ноября 2021 года.
- ↑ Mark S. Eller, Adam Asarch, Barbara A. Gilchrest. Photoprotection in Human Skin—A Multifaceted SOS Response† (англ.) // Photochemistry and Photobiology. — 2008. — Vol. 84, iss. 2. — P. 339–349. — ISSN 1751-1097. — doi:10.1111/j.1751-1097.2007.00264.x. Архивировано 24 ноября 2021 года.
- ↑ С. Г. Инге-Вечтомов. Генетика с основами селекции. — Москва: Высшая школа, 1989
- ↑ А. С. Коничев, Г. А. Севастьянова. Молекулярная биология. — М.: Академия, 2003. — ISBN 5-7695-0783-7.
- ↑ Vilenchik M. M., Knudson A. G. Inverse radiation dose-rate effects on somatic and germ-line mutations and DNA damage rates (англ.) // Proceedings of the National Academy of Sciences. — 2000. — 2 May (vol. 97, no. 10). — P. 5381—5386. — ISSN 0027-8424. — doi:10.1073/pnas.090099497. — PMID 10792040. [исправить]
- ↑ Б. Льюин. Гены.
См. также
- Радиационно-индуцированные фокусы
- Повреждение ДНК
- Клеточный цикл
- Генотерапия
- Первичные радиобиологические процессы
- Прогерия

DNA damage resulting in multiple broken chromosomes
DNA repair is a collection of processes by which a cell identifies and corrects damage to the DNA molecules that encode its genome.[1] In human cells, both normal metabolic activities and environmental factors such as radiation can cause DNA damage, resulting in tens of thousands of individual molecular lesions per cell per day.[2] Many of these lesions cause structural damage to the DNA molecule and can alter or eliminate the cell’s ability to transcribe the gene that the affected DNA encodes. Other lesions induce potentially harmful mutations in the cell’s genome, which affect the survival of its daughter cells after it undergoes mitosis. As a consequence, the DNA repair process is constantly active as it responds to damage in the DNA structure. When normal repair processes fail, and when cellular apoptosis does not occur, irreparable DNA damage may occur, including double-strand breaks and DNA crosslinkages (interstrand crosslinks or ICLs).[3][4] This can eventually lead to malignant tumors, or cancer as per the two hit hypothesis.
The rate of DNA repair is dependent on many factors, including the cell type, the age of the cell, and the extracellular environment. A cell that has accumulated a large amount of DNA damage, or one that no longer effectively repairs damage incurred to its DNA, can enter one of three possible states:
- an irreversible state of dormancy, known as senescence
- cell suicide, also known as apoptosis or programmed cell death
- unregulated cell division, which can lead to the formation of a tumor that is cancerous
The DNA repair ability of a cell is vital to the integrity of its genome and thus to the normal functionality of that organism. Many genes that were initially shown to influence life span have turned out to be involved in DNA damage repair and protection.[5]
Paul Modrich talks about himself and his work in DNA repair.
The 2015 Nobel Prize in Chemistry was awarded to Tomas Lindahl, Paul Modrich, and Aziz Sancar for their work on the molecular mechanisms of DNA repair processes.[6][7]
DNA damage[edit]
DNA damage, due to environmental factors and normal metabolic processes inside the cell, occurs at a rate of 10,000 to 1,000,000 molecular lesions per cell per day.[2] While this constitutes only 0.000165% of the human genome’s approximately 6 billion bases, unrepaired lesions in critical genes (such as tumor suppressor genes) can impede a cell’s ability to carry out its function and appreciably increase the likelihood of tumor formation and contribute to tumour heterogeneity.
The vast majority of DNA damage affects the primary structure of the double helix; that is, the bases themselves are chemically modified. These modifications can in turn disrupt the molecules’ regular helical structure by introducing non-native chemical bonds or bulky adducts that do not fit in the standard double helix. Unlike proteins and RNA, DNA usually lacks tertiary structure and therefore damage or disturbance does not occur at that level. DNA is, however, supercoiled and wound around «packaging» proteins called histones (in eukaryotes), and both superstructures are vulnerable to the effects of DNA damage.
Sources[edit]
DNA damage can be subdivided into two main types:
- endogenous damage such as attack by reactive oxygen species produced from normal metabolic byproducts (spontaneous mutation), especially the process of oxidative deamination
- also includes replication errors
- exogenous damage caused by external agents such as
- ultraviolet [UV 200–400 nm] radiation from the sun or other artificial light sources
- other radiation frequencies, including x-rays and gamma rays
- hydrolysis or thermal disruption
- certain plant toxins
- human-made mutagenic chemicals, especially aromatic compounds that act as DNA intercalating agents
- viruses[8]
The replication of damaged DNA before cell division can lead to the incorporation of wrong bases opposite damaged ones. Daughter cells that inherit these wrong bases carry mutations from which the original DNA sequence is unrecoverable (except in the rare case of a back mutation, for example, through gene conversion).
Types[edit]
There are several types of damage to DNA due to endogenous cellular processes:
- oxidation of bases [e.g. 8-oxo-7,8-dihydroguanine (8-oxoG)] and generation of DNA strand interruptions from reactive oxygen species,
- alkylation of bases (usually methylation), such as formation of 7-methylguanosine, 1-methyladenine, 6-O-Methylguanine
- hydrolysis of bases, such as deamination, depurination, and depyrimidination.
- «bulky adduct formation» (e.g., benzo[a]pyrene diol epoxide-dG adduct, aristolactam I-dA adduct)
- mismatch of bases, due to errors in DNA replication, in which the wrong DNA base is stitched into place in a newly forming DNA strand, or a DNA base is skipped over or mistakenly inserted.
- Monoadduct damage cause by change in single nitrogenous base of DNA
- Diadduct damage
Damage caused by exogenous agents comes in many forms. Some examples are:
- UV-B light causes crosslinking between adjacent cytosine and thymine bases creating pyrimidine dimers. This is called direct DNA damage.
- UV-A light creates mostly free radicals. The damage caused by free radicals is called indirect DNA damage.
- Ionizing radiation such as that created by radioactive decay or in cosmic rays causes breaks in DNA strands. Intermediate-level ionizing radiation may induce irreparable DNA damage (leading to replicational and transcriptional errors needed for neoplasia or may trigger viral interactions) leading to pre-mature aging and cancer.
- Thermal disruption at elevated temperature increases the rate of depurination (loss of purine bases from the DNA backbone) and single-strand breaks. For example, hydrolytic depurination is seen in the thermophilic bacteria, which grow in hot springs at 40–80 °C.[9][10] The rate of depurination (300 purine residues per genome per generation) is too high in these species to be repaired by normal repair machinery, hence a possibility of an adaptive response cannot be ruled out.
- Industrial chemicals such as vinyl chloride and hydrogen peroxide, and environmental chemicals such as polycyclic aromatic hydrocarbons found in smoke, soot and tar create a huge diversity of DNA adducts- ethenobases, oxidized bases, alkylated phosphotriesters and crosslinking of DNA, just to name a few.
UV damage, alkylation/methylation, X-ray damage and oxidative damage are examples of induced damage. Spontaneous damage can include the loss of a base, deamination, sugar ring puckering and tautomeric shift. Constitutive (spontaneous) DNA damage caused by endogenous oxidants can be detected as a low level of histone H2AX phosphorylation in untreated cells.[11]
Nuclear versus mitochondrial[edit]
In human cells, and eukaryotic cells in general, DNA is found in two cellular locations – inside the nucleus and inside the mitochondria. Nuclear DNA (nDNA) exists as chromatin during non-replicative stages of the cell cycle and is condensed into aggregate structures known as chromosomes during cell division. In either state the DNA is highly compacted and wound up around bead-like proteins called histones. Whenever a cell needs to express the genetic information encoded in its nDNA the required chromosomal region is unravelled, genes located therein are expressed, and then the region is condensed back to its resting conformation. Mitochondrial DNA (mtDNA) is located inside mitochondria organelles, exists in multiple copies, and is also tightly associated with a number of proteins to form a complex known as the nucleoid. Inside mitochondria, reactive oxygen species (ROS), or free radicals, byproducts of the constant production of adenosine triphosphate (ATP) via oxidative phosphorylation, create a highly oxidative environment that is known to damage mtDNA. A critical enzyme in counteracting the toxicity of these species is superoxide dismutase, which is present in both the mitochondria and cytoplasm of eukaryotic cells.
Senescence and apoptosis[edit]
Senescence, an irreversible process in which the cell no longer divides, is a protective response to the shortening of the chromosome ends, called telomeres. The telomeres are long regions of repetitive noncoding DNA that cap chromosomes and undergo partial degradation each time a cell undergoes division (see Hayflick limit).[12] In contrast, quiescence is a reversible state of cellular dormancy that is unrelated to genome damage (see cell cycle). Senescence in cells may serve as a functional alternative to apoptosis in cases where the physical presence of a cell for spatial reasons is required by the organism,[13] which serves as a «last resort» mechanism to prevent a cell with damaged DNA from replicating inappropriately in the absence of pro-growth cellular signaling. Unregulated cell division can lead to the formation of a tumor (see cancer), which is potentially lethal to an organism. Therefore, the induction of senescence and apoptosis is considered to be part of a strategy of protection against cancer.[14]
Mutation[edit]
It is important to distinguish between DNA damage and mutation, the two major types of error in DNA. DNA damage and mutation are fundamentally different. Damage results in physical abnormalities in the DNA, such as single- and double-strand breaks, 8-hydroxydeoxyguanosine residues, and polycyclic aromatic hydrocarbon adducts. DNA damage can be recognized by enzymes, and thus can be correctly repaired if redundant information, such as the undamaged sequence in the complementary DNA strand or in a homologous chromosome, is available for copying. If a cell retains DNA damage, transcription of a gene can be prevented, and thus translation into a protein will also be blocked. Replication may also be blocked or the cell may die.
In contrast to DNA damage, a mutation is a change in the base sequence of the DNA. A mutation cannot be recognized by enzymes once the base change is present in both DNA strands, and thus a mutation cannot be repaired. At the cellular level, mutations can cause alterations in protein function and regulation. Mutations are replicated when the cell replicates. In a population of cells, mutant cells will increase or decrease in frequency according to the effects of the mutation on the ability of the cell to survive and reproduce.
Although distinctly different from each other, DNA damage and mutation are related because DNA damage often causes errors of DNA synthesis during replication or repair; these errors are a major source of mutation.
Given these properties of DNA damage and mutation, it can be seen that DNA damage is a special problem in non-dividing or slowly-dividing cells, where unrepaired damage will tend to accumulate over time. On the other hand, in rapidly dividing cells, unrepaired DNA damage that does not kill the cell by blocking replication will tend to cause replication errors and thus mutation. The great majority of mutations that are not neutral in their effect are deleterious to a cell’s survival. Thus, in a population of cells composing a tissue with replicating cells, mutant cells will tend to be lost. However, infrequent mutations that provide a survival advantage will tend to clonally expand at the expense of neighboring cells in the tissue. This advantage to the cell is disadvantageous to the whole organism because such mutant cells can give rise to cancer. Thus, DNA damage in frequently dividing cells, because it gives rise to mutations, is a prominent cause of cancer. In contrast, DNA damage in infrequently-dividing cells is likely a prominent cause of aging.[15]
Mechanisms[edit]
Cells cannot function if DNA damage corrupts the integrity and accessibility of essential information in the genome (but cells remain superficially functional when non-essential genes are missing or damaged). Depending on the type of damage inflicted on the DNA’s double helical structure, a variety of repair strategies have evolved to restore lost information. If possible, cells use the unmodified complementary strand of the DNA or the sister chromatid as a template to recover the original information. Without access to a template, cells use an error-prone recovery mechanism known as translesion synthesis as a last resort.
Damage to DNA alters the spatial configuration of the helix, and such alterations can be detected by the cell. Once damage is localized, specific DNA repair molecules bind at or near the site of damage, inducing other molecules to bind and form a complex that enables the actual repair to take place.
Direct reversal[edit]
Cells are known to eliminate three types of damage to their DNA by chemically reversing it. These mechanisms do not require a template, since the types of damage they counteract can occur in only one of the four bases. Such direct reversal mechanisms are specific to the type of damage incurred and do not involve breakage of the phosphodiester backbone. The formation of pyrimidine dimers upon irradiation with UV light results in an abnormal covalent bond between adjacent pyrimidine bases. The photoreactivation process directly reverses this damage by the action of the enzyme photolyase, whose activation is obligately dependent on energy absorbed from blue/UV light (300–500 nm wavelength) to promote catalysis.[16] Photolyase, an old enzyme present in bacteria, fungi, and most animals no longer functions in humans,[17] who instead use nucleotide excision repair to repair damage from UV irradiation. Another type of damage, methylation of guanine bases, is directly reversed by the enzyme methyl guanine methyl transferase (MGMT), the bacterial equivalent of which is called ogt. This is an expensive process because each MGMT molecule can be used only once; that is, the reaction is stoichiometric rather than catalytic.[18] A generalized response to methylating agents in bacteria is known as the adaptive response and confers a level of resistance to alkylating agents upon sustained exposure by upregulation of alkylation repair enzymes.[19] The third type of DNA damage reversed by cells is certain methylation of the bases cytosine and adenine.
Single-strand damage[edit]

Structure of the base-excision repair enzyme uracil-DNA glycosylase excising a hydrolytically-produced uracil residue from DNA. The uracil residue is shown in yellow.
When only one of the two strands of a double helix has a defect, the other strand can be used as a template to guide the correction of the damaged strand. In order to repair damage to one of the two paired molecules of DNA, there exist a number of excision repair mechanisms that remove the damaged nucleotide and replace it with an undamaged nucleotide complementary to that found in the undamaged DNA strand.[18]
- Base excision repair (BER): damaged single bases or nucleotides are most commonly repaired by removing the base or the nucleotide involved and then inserting the correct base or nucleotide. In base excision repair, a glycosylase[20] enzyme removes the damaged base from the DNA by cleaving the bond between the base and the deoxyribose. These enzymes remove a single base to create an apurinic or apyrimidinic site (AP site).[20] Enzymes called AP endonucleases nick the damaged DNA backbone at the AP site. DNA polymerase then removes the damaged region using its 5’ to 3’ exonuclease activity and correctly synthesizes the new strand using the complementary strand as a template.[20] The gap is then sealed by enzyme DNA ligase.[21]
- Nucleotide excision repair (NER): bulky, helix-distorting damage, such as pyrimidine dimerization caused by UV light is usually repaired by a three-step process. First the damage is recognized, then 12-24 nucleotide-long strands of DNA are removed both upstream and downstream of the damage site by endonucleases, and the removed DNA region is then resynthesized.[22] NER is a highly evolutionarily conserved repair mechanism and is used in nearly all eukaryotic and prokaryotic cells.[22] In prokaryotes, NER is mediated by Uvr proteins.[22] In eukaryotes, many more proteins are involved, although the general strategy is the same.[22]
- Mismatch repair systems are present in essentially all cells to correct errors that are not corrected by proofreading. These systems consist of at least two proteins. One detects the mismatch, and the other recruits an endonuclease that cleaves the newly synthesized DNA strand close to the region of damage. In E. coli , the proteins involved are the Mut class proteins: MutS, MutL, and MutH. In most Eukaryotes, the analog for MutS is MSH and the analog for MutL is MLH. MutH is only present in bacteria. This is followed by removal of damaged region by an exonuclease, resynthesis by DNA polymerase, and nick sealing by DNA ligase.[23]
Double-strand breaks[edit]
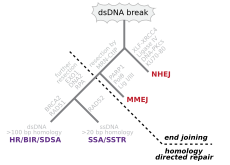
The main double-strand break repair pathways
Double-strand breaks, in which both strands in the double helix are severed, are particularly hazardous to the cell because they can lead to genome rearrangements. In fact, when a double-strand break is accompanied by a cross-linkage joining the two strands at the same point, neither strand can be used as a template for the repair mechanisms, so that the cell will not be able to complete mitosis when it next divides, and will either die or, in rare cases, undergo a mutation.[3][4] Three mechanisms exist to repair double-strand breaks (DSBs): non-homologous end joining (NHEJ), microhomology-mediated end joining (MMEJ), and homologous recombination (HR):[18][24]

DNA ligase, shown above repairing chromosomal damage, is an enzyme that joins broken nucleotides together by catalyzing the formation of an internucleotide ester bond between the phosphate backbone and the deoxyribose nucleotides.
- In NHEJ, DNA Ligase IV, a specialized DNA ligase that forms a complex with the cofactor XRCC4, directly joins the two ends.[25] To guide accurate repair, NHEJ relies on short homologous sequences called microhomologies present on the single-stranded tails of the DNA ends to be joined. If these overhangs are compatible, repair is usually accurate.[26][27][28][29] NHEJ can also introduce mutations during repair. Loss of damaged nucleotides at the break site can lead to deletions, and joining of nonmatching termini forms insertions or translocations. NHEJ is especially important before the cell has replicated its DNA, since there is no template available for repair by homologous recombination. There are «backup» NHEJ pathways in higher eukaryotes.[30] Besides its role as a genome caretaker, NHEJ is required for joining hairpin-capped double-strand breaks induced during V(D)J recombination, the process that generates diversity in B-cell and T-cell receptors in the vertebrate immune system.[31]
- MMEJ starts with short-range end resection by MRE11 nuclease on either side of a double-strand break to reveal microhomology regions.[32] In further steps,[33] Poly (ADP-ribose) polymerase 1 (PARP1) is required and may be an early step in MMEJ. There is pairing of microhomology regions followed by recruitment of flap structure-specific endonuclease 1 (FEN1) to remove overhanging flaps. This is followed by recruitment of XRCC1–LIG3 to the site for ligating the DNA ends, leading to an intact DNA. MMEJ is always accompanied by a deletion, so that MMEJ is a mutagenic pathway for DNA repair.[34]
- HR requires the presence of an identical or nearly identical sequence to be used as a template for repair of the break. The enzymatic machinery responsible for this repair process is nearly identical to the machinery responsible for chromosomal crossover during meiosis. This pathway allows a damaged chromosome to be repaired using a sister chromatid (available in G2 after DNA replication) or a homologous chromosome as a template. DSBs caused by the replication machinery attempting to synthesize across a single-strand break or unrepaired lesion cause collapse of the replication fork and are typically repaired by recombination.
In an in vitro system, MMEJ occurred in mammalian cells at the levels of 10–20% of HR when both HR and NHEJ mechanisms were also available.[32]
The extremophile Deinococcus radiodurans has a remarkable ability to survive DNA damage from ionizing radiation and other sources. At least two copies of the genome, with random DNA breaks, can form DNA fragments through annealing. Partially overlapping fragments are then used for synthesis of homologous regions through a moving D-loop that can continue extension until complementary partner strands are found. In the final step, there is crossover by means of RecA-dependent homologous recombination.[35]
Topoisomerases introduce both single- and double-strand breaks in the course of changing the DNA’s state of supercoiling, which is especially common in regions near an open replication fork. Such breaks are not considered DNA damage because they are a natural intermediate in the topoisomerase biochemical mechanism and are immediately repaired by the enzymes that created them.
Another type of DNA double-strand breaks originates from the DNA heat-sensitive or heat-labile sites. These DNA sites are not initial DSBs. However, they convert to DSB after treating with elevated temperature. Ionizing irradiation can induces a highly complex form of DNA damage as clustered damage. It consists of different types of DNA lesions in various locations of the DNA helix. Some of these closely located lesions can probably convert to DSB by exposure to high temperatures. But the exact nature of these lesions and their interactions is not yet known[36]
Translesion synthesis[edit]
Translesion synthesis (TLS) is a DNA damage tolerance process that allows the DNA replication machinery to replicate past DNA lesions such as thymine dimers or AP sites.[37] It involves switching out regular DNA polymerases for specialized translesion polymerases (i.e. DNA polymerase IV or V, from the Y Polymerase family), often with larger active sites that can facilitate the insertion of bases opposite damaged nucleotides. The polymerase switching is thought to be mediated by, among other factors, the post-translational modification of the replication processivity factor PCNA. Translesion synthesis polymerases often have low fidelity (high propensity to insert wrong bases) on undamaged templates relative to regular polymerases. However, many are extremely efficient at inserting correct bases opposite specific types of damage. For example, Pol η mediates error-free bypass of lesions induced by UV irradiation, whereas Pol ι introduces mutations at these sites. Pol η is known to add the first adenine across the T^T photodimer using Watson-Crick base pairing and the second adenine will be added in its syn conformation using Hoogsteen base pairing. From a cellular perspective, risking the introduction of point mutations during translesion synthesis may be preferable to resorting to more drastic mechanisms of DNA repair, which may cause gross chromosomal aberrations or cell death. In short, the process involves specialized polymerases either bypassing or repairing lesions at locations of stalled DNA replication. For example, Human DNA polymerase eta can bypass complex DNA lesions like guanine-thymine intra-strand crosslink, G[8,5-Me]T, although it can cause targeted and semi-targeted mutations.[38] Paromita Raychaudhury and Ashis Basu[39] studied the toxicity and mutagenesis of the same lesion in Escherichia coli by replicating a G[8,5-Me]T-modified plasmid in E. coli with specific DNA polymerase knockouts. Viability was very low in a strain lacking pol II, pol IV, and pol V, the three SOS-inducible DNA polymerases, indicating that translesion synthesis is conducted primarily by these specialized DNA polymerases.
A bypass platform is provided to these polymerases by Proliferating cell nuclear antigen (PCNA). Under normal circumstances, PCNA bound to polymerases replicates the DNA. At a site of lesion, PCNA is ubiquitinated, or modified, by the RAD6/RAD18 proteins to provide a platform for the specialized polymerases to bypass the lesion and resume DNA replication.[40][41] After translesion synthesis, extension is required. This extension can be carried out by a replicative polymerase if the TLS is error-free, as in the case of Pol η, yet if TLS results in a mismatch, a specialized polymerase is needed to extend it; Pol ζ. Pol ζ is unique in that it can extend terminal mismatches, whereas more processive polymerases cannot. So when a lesion is encountered, the replication fork will stall, PCNA will switch from a processive polymerase to a TLS polymerase such as Pol ι to fix the lesion, then PCNA may switch to Pol ζ to extend the mismatch, and last PCNA will switch to the processive polymerase to continue replication.
Global response to DNA damage[edit]
Cells exposed to ionizing radiation, ultraviolet light or chemicals are prone to acquire multiple sites of bulky DNA lesions and double-strand breaks. Moreover, DNA damaging agents can damage other biomolecules such as proteins, carbohydrates, lipids, and RNA. The accumulation of damage, to be specific, double-strand breaks or adducts stalling the replication forks, are among known stimulation signals for a global response to DNA damage.[42] The global response to damage is an act directed toward the cells’ own preservation and triggers multiple pathways of macromolecular repair, lesion bypass, tolerance, or apoptosis. The common features of global response are induction of multiple genes, cell cycle arrest, and inhibition of cell division.
Initial steps[edit]
The packaging of eukaryotic DNA into chromatin presents a barrier to all DNA-based processes that require recruitment of enzymes to their sites of action. To allow DNA repair, the chromatin must be remodeled. In eukaryotes, ATP dependent chromatin remodeling complexes and histone-modifying enzymes are two predominant factors employed to accomplish this remodeling process.[43]
Chromatin relaxation occurs rapidly at the site of a DNA damage.[44][45] In one of the earliest steps, the stress-activated protein kinase, c-Jun N-terminal kinase (JNK), phosphorylates SIRT6 on serine 10 in response to double-strand breaks or other DNA damage.[46] This post-translational modification facilitates the mobilization of SIRT6 to DNA damage sites, and is required for efficient recruitment of poly (ADP-ribose) polymerase 1 (PARP1) to DNA break sites and for efficient repair of DSBs.[46] PARP1 protein starts to appear at DNA damage sites in less than a second, with half maximum accumulation within 1.6 seconds after the damage occurs.[47] PARP1 synthesizes polymeric adenosine diphosphate ribose (poly (ADP-ribose) or PAR) chains on itself. Next the chromatin remodeler ALC1 quickly attaches to the product of PARP1 action, a poly-ADP ribose chain, and ALC1 completes arrival at the DNA damage within 10 seconds of the occurrence of the damage.[45] About half of the maximum chromatin relaxation, presumably due to action of ALC1, occurs by 10 seconds.[45] This then allows recruitment of the DNA repair enzyme MRE11, to initiate DNA repair, within 13 seconds.[47]
γH2AX, the phosphorylated form of H2AX is also involved in the early steps leading to chromatin decondensation after DNA double-strand breaks. The histone variant H2AX constitutes about 10% of the H2A histones in human chromatin.[48] γH2AX (H2AX phosphorylated on serine 139) can be detected as soon as 20 seconds after irradiation of cells (with DNA double-strand break formation), and half maximum accumulation of γH2AX occurs in one minute.[48] The extent of chromatin with phosphorylated γH2AX is about two million base pairs at the site of a DNA double-strand break.[48] γH2AX does not, itself, cause chromatin decondensation, but within 30 seconds of irradiation, RNF8 protein can be detected in association with γH2AX.[49] RNF8 mediates extensive chromatin decondensation, through its subsequent interaction with CHD4,[50] a component of the nucleosome remodeling and deacetylase complex NuRD.
DDB2 occurs in a heterodimeric complex with DDB1. This complex further complexes with the ubiquitin ligase protein CUL4A[51] and with PARP1.[52] This larger complex rapidly associates with UV-induced damage within chromatin, with half-maximum association completed in 40 seconds.[51] The PARP1 protein, attached to both DDB1 and DDB2, then PARylates (creates a poly-ADP ribose chain) on DDB2 that attracts the DNA remodeling protein ALC1.[52] Action of ALC1 relaxes the chromatin at the site of UV damage to DNA. This relaxation allows other proteins in the nucleotide excision repair pathway to enter the chromatin and repair UV-induced cyclobutane pyrimidine dimer damages.
After rapid chromatin remodeling, cell cycle checkpoints are activated to allow DNA repair to occur before the cell cycle progresses. First, two kinases, ATM and ATR are activated within 5 or 6 minutes after DNA is damaged. This is followed by phosphorylation of the cell cycle checkpoint protein Chk1, initiating its function, about 10 minutes after DNA is damaged.[53]
DNA damage checkpoints[edit]
After DNA damage, cell cycle checkpoints are activated. Checkpoint activation pauses the cell cycle and gives the cell time to repair the damage before continuing to divide. DNA damage checkpoints occur at the G1/S and G2/M boundaries. An intra-S checkpoint also exists. Checkpoint activation is controlled by two master kinases, ATM and ATR. ATM responds to DNA double-strand breaks and disruptions in chromatin structure,[54] whereas ATR primarily responds to stalled replication forks. These kinases phosphorylate downstream targets in a signal transduction cascade, eventually leading to cell cycle arrest. A class of checkpoint mediator proteins including BRCA1, MDC1, and 53BP1 has also been identified.[55] These proteins seem to be required for transmitting the checkpoint activation signal to downstream proteins.
DNA damage checkpoint is a signal transduction pathway that blocks cell cycle progression in G1, G2 and metaphase and slows down the rate of S phase progression when DNA is damaged. It leads to a pause in cell cycle allowing the cell time to repair the damage before continuing to divide.
Checkpoint Proteins can be separated into four groups: phosphatidylinositol 3-kinase (PI3K)-like protein kinase, proliferating cell nuclear antigen (PCNA)-like group, two serine/threonine(S/T) kinases and their adaptors. Central to all DNA damage induced checkpoints responses is a pair of large protein kinases belonging to the first group of PI3K-like protein kinases-the ATM (Ataxia telangiectasia mutated) and ATR (Ataxia- and Rad-related) kinases, whose sequence and functions have been well conserved in evolution. All DNA damage response requires either ATM or ATR because they have the ability to bind to the chromosomes at the site of DNA damage, together with accessory proteins that are platforms on which DNA damage response components and DNA repair complexes can be assembled.
An important downstream target of ATM and ATR is p53, as it is required for inducing apoptosis following DNA damage.[56] The cyclin-dependent kinase inhibitor p21 is induced by both p53-dependent and p53-independent mechanisms and can arrest the cell cycle at the G1/S and G2/M checkpoints by deactivating cyclin/cyclin-dependent kinase complexes.[57]
The prokaryotic SOS response[edit]
The SOS response is the changes in gene expression in Escherichia coli and other bacteria in response to extensive DNA damage. The prokaryotic SOS system is regulated by two key proteins: LexA and RecA. The LexA homodimer is a transcriptional repressor that binds to operator sequences commonly referred to as SOS boxes. In Escherichia coli it is known that LexA regulates transcription of approximately 48 genes including the lexA and recA genes.[58] The SOS response is known to be widespread in the Bacteria domain, but it is mostly absent in some bacterial phyla, like the Spirochetes.[59]
The most common cellular signals activating the SOS response are regions of single-stranded DNA (ssDNA), arising from stalled replication forks or double-strand breaks, which are processed by DNA helicase to separate the two DNA strands.[42] In the initiation step, RecA protein binds to ssDNA in an ATP hydrolysis driven reaction creating RecA–ssDNA filaments. RecA–ssDNA filaments activate LexA autoprotease activity, which ultimately leads to cleavage of LexA dimer and subsequent LexA degradation. The loss of LexA repressor induces transcription of the SOS genes and allows for further signal induction, inhibition of cell division and an increase in levels of proteins responsible for damage processing.
In Escherichia coli, SOS boxes are 20-nucleotide long sequences near promoters with palindromic structure and a high degree of sequence conservation. In other classes and phyla, the sequence of SOS boxes varies considerably, with different length and composition, but it is always highly conserved and one of the strongest short signals in the genome.[59] The high information content of SOS boxes permits differential binding of LexA to different promoters and allows for timing of the SOS response. The lesion repair genes are induced at the beginning of SOS response. The error-prone translesion polymerases, for example, UmuCD’2 (also called DNA polymerase V), are induced later on as a last resort.[60] Once the DNA damage is repaired or bypassed using polymerases or through recombination, the amount of single-stranded DNA in cells is decreased, lowering the amounts of RecA filaments decreases cleavage activity of LexA homodimer, which then binds to the SOS boxes near promoters and restores normal gene expression.
Eukaryotic transcriptional responses to DNA damage[edit]
Eukaryotic cells exposed to DNA damaging agents also activate important defensive pathways by inducing multiple proteins involved in DNA repair, cell cycle checkpoint control, protein trafficking and degradation. Such genome wide transcriptional response is very complex and tightly regulated, thus allowing coordinated global response to damage. Exposure of yeast Saccharomyces cerevisiae to DNA damaging agents results in overlapping but distinct transcriptional profiles. Similarities to environmental shock response indicates that a general global stress response pathway exist at the level of transcriptional activation. In contrast, different human cell types respond to damage differently indicating an absence of a common global response. The probable explanation for this difference between yeast and human cells may be in the heterogeneity of mammalian cells. In an animal different types of cells are distributed among different organs that have evolved different sensitivities to DNA damage.[61]
In general global response to DNA damage involves expression of multiple genes responsible for postreplication repair, homologous recombination, nucleotide excision repair, DNA damage checkpoint, global transcriptional activation, genes controlling mRNA decay, and many others. A large amount of damage to a cell leaves it with an important decision: undergo apoptosis and die, or survive at the cost of living with a modified genome. An increase in tolerance to damage can lead to an increased rate of survival that will allow a greater accumulation of mutations. Yeast Rev1 and human polymerase η are members of Y family translesion DNA polymerases present during global response to DNA damage and are responsible for enhanced mutagenesis during a global response to DNA damage in eukaryotes.[42]
Aging[edit]
Pathological effects of poor DNA repair[edit]

DNA repair rate is an important determinant of cell pathology.
Experimental animals with genetic deficiencies in DNA repair often show decreased life span and increased cancer incidence.[15] For example, mice deficient in the dominant NHEJ pathway and in telomere maintenance mechanisms get lymphoma and infections more often, and, as a consequence, have shorter lifespans than wild-type mice.[62] In similar manner, mice deficient in a key repair and transcription protein that unwinds DNA helices have premature onset of aging-related diseases and consequent shortening of lifespan.[63] However, not every DNA repair deficiency creates exactly the predicted effects; mice deficient in the NER pathway exhibited shortened life span without correspondingly higher rates of mutation.[64]
If the rate of DNA damage exceeds the capacity of the cell to repair it, the accumulation of errors can overwhelm the cell and result in early senescence, apoptosis, or cancer. Inherited diseases associated with faulty DNA repair functioning result in premature aging,[15] increased sensitivity to carcinogens and correspondingly increased cancer risk (see below). On the other hand, organisms with enhanced DNA repair systems, such as Deinococcus radiodurans, the most radiation-resistant known organism, exhibit remarkable resistance to the double-strand break-inducing effects of radioactivity, likely due to enhanced efficiency of DNA repair and especially NHEJ.[65]
Longevity and caloric restriction[edit]

Most life span influencing genes affect the rate of DNA damage.
A number of individual genes have been identified as influencing variations in life span within a population of organisms. The effects of these genes is strongly dependent on the environment, in particular, on the organism’s diet. Caloric restriction reproducibly results in extended lifespan in a variety of organisms, likely via nutrient sensing pathways and decreased metabolic rate. The molecular mechanisms by which such restriction results in lengthened lifespan are as yet unclear (see[66] for some discussion); however, the behavior of many genes known to be involved in DNA repair is altered under conditions of caloric restriction. Several agents reported to have anti-aging properties have been shown to attenuate constitutive level of mTOR signaling, an evidence of reduction of metabolic activity, and concurrently to reduce constitutive level of DNA damage induced by endogenously generated reactive oxygen species.[67]
For example, increasing the gene dosage of the gene SIR-2, which regulates DNA packaging in the nematode worm Caenorhabditis elegans, can significantly extend lifespan.[68] The mammalian homolog of SIR-2 is known to induce downstream DNA repair factors involved in NHEJ, an activity that is especially promoted under conditions of caloric restriction.[69] Caloric restriction has been closely linked to the rate of base excision repair in the nuclear DNA of rodents,[70] although similar effects have not been observed in mitochondrial DNA.[71]
The C. elegans gene AGE-1, an upstream effector of DNA repair pathways, confers dramatically extended life span under free-feeding conditions but leads to a decrease in reproductive fitness under conditions of caloric restriction.[72] This observation supports the pleiotropy theory of the biological origins of aging, which suggests that genes conferring a large survival advantage early in life will be selected for even if they carry a corresponding disadvantage late in life.
Medicine and DNA repair modulation[edit]
Hereditary DNA repair disorders[edit]
Defects in the NER mechanism are responsible for several genetic disorders, including:
- Xeroderma pigmentosum: hypersensitivity to sunlight/UV, resulting in increased skin cancer incidence and premature aging
- Cockayne syndrome: hypersensitivity to UV and chemical agents
- Trichothiodystrophy: sensitive skin, brittle hair and nails
Mental retardation often accompanies the latter two disorders, suggesting increased vulnerability of developmental neurons.
Other DNA repair disorders include:
- Werner’s syndrome: premature aging and retarded growth
- Bloom’s syndrome: sunlight hypersensitivity, high incidence of malignancies (especially leukemias).
- Ataxia telangiectasia: sensitivity to ionizing radiation and some chemical agents
All of the above diseases are often called «segmental progerias» («accelerated aging diseases») because those affected appear elderly and experience aging-related diseases at an abnormally young age, while not manifesting all the symptoms of old age.
Other diseases associated with reduced DNA repair function include Fanconi anemia, hereditary breast cancer and hereditary colon cancer.
Cancer[edit]
Because of inherent limitations in the DNA repair mechanisms, if humans lived long enough, they would all eventually develop cancer.[73][74] There are at least 34 Inherited human DNA repair gene mutations that increase cancer risk. Many of these mutations cause DNA repair to be less effective than normal. In particular, Hereditary nonpolyposis colorectal cancer (HNPCC) is strongly associated with specific mutations in the DNA mismatch repair pathway. BRCA1 and BRCA2, two important genes whose mutations confer a hugely increased risk of breast cancer on carriers,[75] are both associated with a large number of DNA repair pathways, especially NHEJ and homologous recombination.
Cancer therapy procedures such as chemotherapy and radiotherapy work by overwhelming the capacity of the cell to repair DNA damage, resulting in cell death. Cells that are most rapidly dividing – most typically cancer cells – are preferentially affected. The side-effect is that other non-cancerous but rapidly dividing cells such as progenitor cells in the gut, skin, and hematopoietic system are also affected. Modern cancer treatments attempt to localize the DNA damage to cells and tissues only associated with cancer, either by physical means (concentrating the therapeutic agent in the region of the tumor) or by biochemical means (exploiting a feature unique to cancer cells in the body). In the context of therapies targeting DNA damage response genes, the latter approach has been termed ‘synthetic lethality’.[76]
Perhaps the most well-known of these ‘synthetic lethality’ drugs is the poly(ADP-ribose) polymerase 1 (PARP1) inhibitor olaparib, which was approved by the Food and Drug Administration in 2015 for the treatment in women of BRCA-defective ovarian cancer. Tumor cells with partial loss of DNA damage response (specifically, homologous recombination repair) are dependent on another mechanism – single-strand break repair – which is a mechanism consisting, in part, of the PARP1 gene product.[77] Olaparib is combined with chemotherapeutics to inhibit single-strand break repair induced by DNA damage caused by the co-administered chemotherapy. Tumor cells relying on this residual DNA repair mechanism are unable to repair the damage and hence are not able to survive and proliferate, whereas normal cells can repair the damage with the functioning homologous recombination mechanism.
Many other drugs for use against other residual DNA repair mechanisms commonly found in cancer are currently under investigation. However, synthetic lethality therapeutic approaches have been questioned due to emerging evidence of acquired resistance, achieved through rewiring of DNA damage response pathways and reversion of previously inhibited defects.[78]
DNA repair defects in cancer[edit]
It has become apparent over the past several years that the DNA damage response acts as a barrier to the malignant transformation of preneoplastic cells.[79] Previous studies have shown an elevated DNA damage response in cell-culture models with oncogene activation[80] and preneoplastic colon adenomas.[81] DNA damage response mechanisms trigger cell-cycle arrest, and attempt to repair DNA lesions or promote cell death/senescence if repair is not possible. Replication stress is observed in preneoplastic cells due to increased proliferation signals from oncogenic mutations. Replication stress is characterized by: increased replication initiation/origin firing; increased transcription and collisions of transcription-replication complexes; nucleotide deficiency; increase in reactive oxygen species (ROS).[82]
Replication stress, along with the selection for inactivating mutations in DNA damage response genes in the evolution of the tumor,[83] leads to downregulation and/or loss of some DNA damage response mechanisms, and hence loss of DNA repair and/or senescence/programmed cell death. In experimental mouse models, loss of DNA damage response-mediated cell senescence was observed after using a short hairpin RNA (shRNA) to inhibit the double-strand break response kinase ataxia telangiectasia (ATM), leading to increased tumor size and invasiveness.[81] Humans born with inherited defects in DNA repair mechanisms (for example, Li-Fraumeni syndrome) have a higher cancer risk.[84]
The prevalence of DNA damage response mutations differs across cancer types; for example, 30% of breast invasive carcinomas have mutations in genes involved in homologous recombination.[79] In cancer, downregulation is observed across all DNA damage response mechanisms (base excision repair (BER), nucleotide excision repair (NER), DNA mismatch repair (MMR), homologous recombination repair (HR), non-homologous end joining (NHEJ) and translesion DNA synthesis (TLS).[85] As well as mutations to DNA damage repair genes, mutations also arise in the genes responsible for arresting the cell cycle to allow sufficient time for DNA repair to occur, and some genes are involved in both DNA damage repair and cell cycle checkpoint control, for example ATM and checkpoint kinase 2 (CHEK2) – a tumor suppressor that is often absent or downregulated in non-small cell lung cancer.[86]
Genes involved in DNA damage response pathways and frequently mutated in cancer (HR = homologous recombination; NHEJ = non-homologous end joining; SSA = single-strand annealing; FA = fanconi anemia pathway; BER = base excision repair; NER = nucleotide excision repair; MMR = mismatch repair)
| HR | NHEJ | SSA | FA | BER | NER | MMR | |
|---|---|---|---|---|---|---|---|
| ATM | |||||||
| ATR | |||||||
| PAXIP | |||||||
| RPA | |||||||
| BRCA1 | |||||||
| BRCA2 | |||||||
| RAD51 | |||||||
| RFC | |||||||
| XRCC1 | |||||||
| PCNA | |||||||
| PARP1 | |||||||
| ERCC1 | |||||||
| MSH3 |
Epigenetic DNA repair defects in cancer[edit]
Classically, cancer has been viewed as a set of diseases that are driven by progressive genetic abnormalities that include mutations in tumour-suppressor genes and oncogenes, and chromosomal aberrations. However, it has become apparent that cancer is also driven by
epigenetic alterations.[87]
Epigenetic alterations refer to functionally relevant modifications to the genome that do not involve a change in the nucleotide sequence. Examples of such modifications are changes in DNA methylation (hypermethylation and hypomethylation) and histone modification,[88] changes in chromosomal architecture (caused by inappropriate expression of proteins such as HMGA2 or HMGA1)[89] and changes caused by microRNAs. Each of these epigenetic alterations serves to regulate gene expression without altering the underlying DNA sequence. These changes usually remain through cell divisions, last for multiple cell generations, and can be considered to be epimutations (equivalent to mutations).
While large numbers of epigenetic alterations are found in cancers, the epigenetic alterations in DNA repair genes, causing reduced expression of DNA repair proteins, appear to be particularly important. Such alterations are thought to occur early in progression to cancer and to be a likely cause of the genetic instability characteristic of cancers.[90][91][92]
Reduced expression of DNA repair genes causes deficient DNA repair. When DNA repair is deficient DNA damages remain in cells at a higher than usual level and these excess damages cause increased frequencies of mutation or epimutation. Mutation rates increase substantially in cells defective in DNA mismatch repair[93][94] or in homologous recombinational repair (HRR).[95] Chromosomal rearrangements and aneuploidy also increase in HRR defective cells.[96]
Higher levels of DNA damage not only cause increased mutation, but also cause increased epimutation. During repair of DNA double strand breaks, or repair of other DNA damages, incompletely cleared sites of repair can cause epigenetic gene silencing.[97][98]
Deficient expression of DNA repair proteins due to an inherited mutation can cause increased risk of cancer. Individuals with an inherited impairment in any of 34 DNA repair genes (see article DNA repair-deficiency disorder) have an increased risk of cancer, with some defects causing up to a 100% lifetime chance of cancer (e.g. p53 mutations).[99] However, such germline mutations (which cause highly penetrant cancer syndromes) are the cause of only about 1 percent of cancers.[100]
Frequencies of epimutations in DNA repair genes[edit]

A chart of common DNA damaging agents, examples of lesions they cause in DNA, and pathways used to repair these lesions. Also shown are many of the genes in these pathways, an indication of which genes are epigenetically regulated to have reduced (or increased) expression in various cancers. It also shows genes in the error-prone microhomology-mediated end joining pathway with increased expression in various cancers.
Deficiencies in DNA repair enzymes are occasionally caused by a newly arising somatic mutation in a DNA repair gene, but are much more frequently caused by epigenetic alterations that reduce or silence expression of DNA repair genes. For example, when 113 colorectal cancers were examined in sequence, only four had a missense mutation in the DNA repair gene MGMT, while the majority had reduced MGMT expression due to methylation of the MGMT promoter region (an epigenetic alteration).[101] Five different studies found that between 40% and 90% of colorectal cancers have reduced MGMT expression due to methylation of the MGMT promoter region.[102][103][104][105][106]
Similarly, out of 119 cases of mismatch repair-deficient colorectal cancers that lacked DNA repair gene PMS2 expression, PMS2 was deficient in 6 due to mutations in the PMS2 gene, while in 103 cases PMS2 expression was deficient because its pairing partner MLH1 was repressed due to promoter methylation (PMS2 protein is unstable in the absence of MLH1).[107] In the other 10 cases, loss of PMS2 expression was likely due to epigenetic overexpression of the microRNA, miR-155, which down-regulates MLH1.[108]
In a further example, epigenetic defects were found in various cancers (e.g. breast, ovarian, colorectal and head and neck). Two or three deficiencies in the expression of ERCC1, XPF or PMS2 occur simultaneously in the majority of 49 colon cancers evaluated by Facista et al.[109]
The chart in this section shows some frequent DNA damaging agents, examples of DNA lesions they cause, and the pathways that deal with these DNA damages. At least 169 enzymes are either directly employed in DNA repair or influence DNA repair processes.[110] Of these, 83 are directly employed in repairing the 5 types of DNA damages illustrated in the chart.
Some of the more well studied genes central to these repair processes are shown in the chart. The gene designations shown in red, gray or cyan indicate genes frequently epigenetically altered in various types of cancers. Wikipedia articles on each of the genes highlighted by red, gray or cyan describe the epigenetic alteration(s) and the cancer(s) in which these epimutations are found. Review articles,[111] and broad experimental survey articles[112][113] also document most of these epigenetic DNA repair deficiencies in cancers.
Red-highlighted genes are frequently reduced or silenced by epigenetic mechanisms in various cancers. When these genes have low or absent expression, DNA damages can accumulate. Replication errors past these damages (see translesion synthesis) can lead to increased mutations and, ultimately, cancer. Epigenetic repression of DNA repair genes in accurate DNA repair pathways appear to be central to carcinogenesis.
The two gray-highlighted genes RAD51 and BRCA2, are required for homologous recombinational repair. They are sometimes epigenetically over-expressed and sometimes under-expressed in certain cancers. As indicated in the Wikipedia articles on RAD51 and BRCA2, such cancers ordinarily have epigenetic deficiencies in other DNA repair genes. These repair deficiencies would likely cause increased unrepaired DNA damages. The over-expression of RAD51 and BRCA2 seen in these cancers may reflect selective pressures for compensatory RAD51 or BRCA2 over-expression and increased homologous recombinational repair to at least partially deal with such excess DNA damages. In those cases where RAD51 or BRCA2 are under-expressed, this would itself lead to increased unrepaired DNA damages. Replication errors past these damages (see translesion synthesis) could cause increased mutations and cancer, so that under-expression of RAD51 or BRCA2 would be carcinogenic in itself.
Cyan-highlighted genes are in the microhomology-mediated end joining (MMEJ) pathway and are up-regulated in cancer. MMEJ is an additional error-prone inaccurate repair pathway for double-strand breaks. In MMEJ repair of a double-strand break, an homology of 5–25 complementary base pairs between both paired strands is sufficient to align the strands, but mismatched ends (flaps) are usually present. MMEJ removes the extra nucleotides (flaps) where strands are joined, and then ligates the strands to create an intact DNA double helix. MMEJ almost always involves at least a small deletion, so that it is a mutagenic pathway.[24] FEN1, the flap endonuclease in MMEJ, is epigenetically increased by promoter hypomethylation and is over-expressed in the majority of cancers of the breast,[114] prostate,[115] stomach,[116][117] neuroblastomas,[118] pancreas,[119] and lung.[120] PARP1 is also over-expressed when its promoter region ETS site is epigenetically hypomethylated, and this contributes to progression to endometrial cancer[121] and BRCA-mutated serous ovarian cancer.[122] Other genes in the MMEJ pathway are also over-expressed in a number of cancers (see MMEJ for summary), and are also shown in cyan.
Genome-wide distribution of DNA repair in human somatic cells[edit]
Differential activity of DNA repair pathways across various regions of the human genome causes mutations to be very unevenly distributed within tumor genomes.[123][124] In particular, the gene-rich, early-replicating regions of the human genome exhibit lower mutation frequencies than the gene-poor, late-replicating heterochromatin. One mechanism underlying this involves the histone modification H3K36me3, which can recruit mismatch repair proteins,[125] thereby lowering mutation rates in H3K36me3-marked regions.[126] Another important mechanism concerns nucleotide excision repair, which can be recruited by the transcription machinery, lowering somatic mutation rates in active genes[124] and other open chromatin regions.[127]
Epigenetic alterations due to DNA repair[edit]
Damage to DNA is very common and is constantly being repaired. Epigenetic alterations can accompany DNA repair of oxidative damage or double-strand breaks. In human cells, oxidative DNA damage occurs about 10,000 times a day and DNA double-strand breaks occur about 10 to 50 times a cell cycle in somatic replicating cells (see DNA damage (naturally occurring)). The selective advantage of DNA repair is to allow the cell to survive in the face of DNA damage. The selective advantage of epigenetic alterations that occur with DNA repair is not clear.
Repair of oxidative DNA damage can alter epigenetic markers[edit]
In the steady state (with endogenous damages occurring and being repaired), there are about 2,400 oxidatively damaged guanines that form 8-oxo-2′-deoxyguanosine (8-OHdG) in the average mamalian cell DNA.[128] 8-OHdG constitutes about 5% of the oxidative damages commonly present in DNA.[129] The oxidized guanines do not occur randomly among all guanines in DNA. There is a sequence preference for the guanine at a methylated CpG site (a cytosine followed by guanine along its 5′ → 3′ direction and where the cytosine is methylated (5-mCpG)).[130] A 5-mCpG site has the lowest ionization potential for guanine oxidation.

Initiation of DNA demethylation at a CpG site. In adult somatic cells DNA methylation typically occurs in the context of CpG dinucleotides (CpG sites), forming 5-methylcytosine-pG, or 5mCpG. Reactive oxygen species (ROS) may attack guanine at the dinucleotide site, forming 8-hydroxy-2′-deoxyguanosine (8-OHdG), and resulting in a 5mCp-8-OHdG dinucleotide site. The base excision repair enzyme OGG1 targets 8-OHdG and binds to the lesion without immediate excision. OGG1, present at a 5mCp-8-OHdG site recruits TET1 and TET1 oxidizes the 5mC adjacent to the 8-OHdG. This initiates demethylation of 5mC.[131]
Oxidized guanine has mispairing potential and is mutagenic.[132] Oxoguanine glycosylase (OGG1) is the primary enzyme responsible for the excision of the oxidized guanine during DNA repair. OGG1 finds and binds to an 8-OHdG within a few seconds.[133] However, OGG1 does not immediately excise 8-OHdG. In HeLa cells half maximum removal of 8-OHdG occurs in 30 minutes,[134] and in irradiated mice, the 8-OHdGs induced in the mouse liver are removed with a half-life of 11 minutes.[129]
When OGG1 is present at an oxidized guanine within a methylated CpG site it recruits TET1 to the 8-OHdG lesion (see Figure). This allows TET1 to demethylate an adjacent methylated cytosine. Demethylation of cytosine is an epigenetic alteration.
As an example, when human mammary epithelial cells were treated with H2O2 for six hours, 8-OHdG increased about 3.5-fold in DNA and this caused about 80% demethylation of the 5-methylcytosines in the genome.[131] Demethylation of CpGs in a gene promoter by TET enzyme activity increases transcription of the gene into messenger RNA.[135] In cells treated with H2O2, one particular gene was examined, BACE1.[131] The methylation level of the BACE1 CpG island was reduced (an epigenetic alteration) and this allowed about 6.5 fold increase of expression of BACE1 messenger RNA.
While six-hour incubation with H2O2 causes considerable demethylation of 5-mCpG sites, shorter times of H2O2 incubation appear to promote other epigenetic alterations. Treatment of cells with H2O2 for 30 minutes causes the mismatch repair protein heterodimer MSH2-MSH6 to recruit DNA methyltransferase 1 (DNMT1) to sites of some kinds of oxidative DNA damage.[136] This could cause increased methylation of cytosines (epigenetic alterations) at these locations.
Jiang et al.[137] treated HEK 293 cells with agents causing oxidative DNA damage, (potassium bromate (KBrO3) or potassium chromate (K2CrO4)). Base excision repair (BER) of oxidative damage occurred with the DNA repair enzyme polymerase beta localizing to oxidized guanines. Polymerase beta is the main human polymerase in short-patch BER of oxidative DNA damage. Jiang et al.[137] also found that polymerase beta recruited the DNA methyltransferase protein DNMT3b to BER repair sites. They then evaluated the methylation pattern at the single nucleotide level in a small region of DNA including the promoter region and the early transcription region of the BRCA1 gene. Oxidative DNA damage from bromate modulated the DNA methylation pattern (caused epigenetic alterations) at CpG sites within the region of DNA studied. In untreated cells, CpGs located at −189, −134, −29, −19, +16, and +19 of the BRCA1 gene had methylated cytosines (where numbering is from the messenger RNA transcription start site, and negative numbers indicate nucleotides in the upstream promoter region). Bromate treatment-induced oxidation resulted in the loss of cytosine methylation at −189, −134, +16 and +19 while also leading to the formation of new methylation at the CpGs located at −80, −55, −21 and +8 after DNA repair was allowed.
Homologous recombinational repair alters epigenetic markers[edit]
At least four articles report the recruitment of DNA methyltransferase 1 (DNMT1) to sites of DNA double-strand breaks.[138][139][140][141] During homologous recombinational repair (HR) of the double-strand break, the involvement of DNMT1 causes the two repaired strands of DNA to have different levels of methylated cytosines. One strand becomes frequently methylated at about 21 CpG sites downstream of the repaired double-strand break. The other DNA strand loses methylation at about six CpG sites that were previously methylated downstream of the double-strand break, as well as losing methylation at about five CpG sites that were previously methylated upstream of the double-strand break. When the chromosome is replicated, this gives rise to one daughter chromosome that is heavily methylated downstream of the previous break site and one that is unmethylated in the region both upstream and downstream of the previous break site. With respect to the gene that was broken by the double-strand break, half of the progeny cells express that gene at a high level and in the other half of the progeny cells expression of that gene is repressed. When clones of these cells were maintained for three years, the new methylation patterns were maintained over that time period.[142]
In mice with a CRISPR-mediated homology-directed recombination insertion in their genome there were a large number of increased methylations of CpG sites within the double-strand break-associated insertion.[143]
Non-homologous end joining can cause some epigenetic marker alterations[edit]
Non-homologous end joining (NHEJ) repair of a double-strand break can cause a small number of demethylations of pre-existing cytosine DNA methylations downstream of the repaired double-strand break.[139] Further work by Allen et al.[144] showed that NHEJ of a DNA double-strand break in a cell could give rise to some progeny cells having repressed expression of the gene harboring the initial double-strand break and some progeny having high expression of that gene due to epigenetic alterations associated with NHEJ repair. The frequency of epigenetic alterations causing repression of a gene after an NHEJ repair of a DNA double-strand break in that gene may be about 0.9%.[140]
Evolution[edit]
The basic processes of DNA repair are highly conserved among both prokaryotes and eukaryotes and even among bacteriophages (viruses which infect bacteria); however, more complex organisms with more complex genomes have correspondingly more complex repair mechanisms.[145] The ability of a large number of protein structural motifs to catalyze relevant chemical reactions has played a significant role in the elaboration of repair mechanisms during evolution. For an extremely detailed review of hypotheses relating to the evolution of DNA repair, see.[146]
The fossil record indicates that single-cell life began to proliferate on the planet at some point during the Precambrian period, although exactly when recognizably modern life first emerged is unclear. Nucleic acids became the sole and universal means of encoding genetic information, requiring DNA repair mechanisms that in their basic form have been inherited by all extant life forms from their common ancestor. The emergence of Earth’s oxygen-rich atmosphere (known as the «oxygen catastrophe») due to photosynthetic organisms, as well as the presence of potentially damaging free radicals in the cell due to oxidative phosphorylation, necessitated the evolution of DNA repair mechanisms that act specifically to counter the types of damage induced by oxidative stress.
Rate of evolutionary change[edit]
On some occasions, DNA damage is not repaired or is repaired by an error-prone mechanism that results in a change from the original sequence. When this occurs, mutations may propagate into the genomes of the cell’s progeny. Should such an event occur in a germ line cell that will eventually produce a gamete, the mutation has the potential to be passed on to the organism’s offspring. The rate of evolution in a particular species (or, in a particular gene) is a function of the rate of mutation. As a consequence, the rate and accuracy of DNA repair mechanisms have an influence over the process of evolutionary change.[147] DNA damage protection and repair does not influence the rate of adaptation by gene regulation and by recombination and selection of alleles. On the other hand, DNA damage repair and protection does influence the rate of accumulation of irreparable, advantageous, code expanding, inheritable mutations, and slows down the evolutionary mechanism for expansion of the genome of organisms with new functionalities. The tension between evolvability and mutation repair and protection needs further investigation.
Technology[edit]
A technology named clustered regularly interspaced short palindromic repeat (shortened to CRISPR-Cas9) was discovered in 2012. The new technology allows anyone with molecular biology training to alter the genes of any species with precision, by inducing DNA damage at a specific point and then altering DNA repair mechanisms to insert new genes.[148] It is cheaper, more efficient, and more precise than other technologies. With the help of CRISPR–Cas9, parts of a genome can be edited by scientists by removing, adding, or altering parts in a DNA sequence.
See also[edit]
- Accelerated aging disease
- Aging DNA
- Cell cycle
- DNA damage (naturally occurring)
- DNA damage theory of aging
- DNA replication
- Direct DNA damage
- Gene therapy
- Human mitochondrial genetics
- Indirect DNA damage
- Life extension
- Progeria
- REPAIRtoire
- Senescence
- SiDNA
- The scientific journal DNA Repair under Mutation Research
References[edit]
- ^ «Nature Reviews Series: DNA damage». Nature Reviews Molecular Cell Biology. 5 July 2017. Retrieved 7 November 2018.
- ^ a b Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J (2004). Molecular Biology of the Cell (5th ed.). New York: WH Freeman. p. 963.
- ^ a b Acharya PV (1971). «The isolation and partial characterization of age-correlated oligo-deoxyribo-ribonucleotides with covalently linked aspartyl-glutamyl polypeptides». Johns Hopkins Medical Journal. Supplement (1): 254–60. PMID 5055816.
- ^ a b Bjorksten J, Acharya PV, Ashman S, Wetlaufer DB (July 1971). «Gerogenic fractions in the tritiated rat». Journal of the American Geriatrics Society. 19 (7): 561–74. doi:10.1111/j.1532-5415.1971.tb02577.x. PMID 5106728. S2CID 33154242.
- ^ Browner WS, Kahn AJ, Ziv E, Reiner AP, Oshima J, Cawthon RM, et al. (December 2004). «The genetics of human longevity». The American Journal of Medicine. 117 (11): 851–60. CiteSeerX 10.1.1.556.6874. doi:10.1016/j.amjmed.2004.06.033. PMID 15589490.
- ^ Broad WJ (7 October 2015). «Nobel Prize in Chemistry Awarded to Tomas Lindahl, Paul Modrich and Aziz Sancar for DNA Studies». The New York Times. Retrieved 7 October 2015.
- ^ Staff (7 October 2015). «The Nobel Prize in Chemistry 2015 – DNA repair – providing chemical stability for life» (PDF). Nobel Prize. Retrieved 7 October 2015.
- ^ Roulston A, Marcellus RC, Branton PE (1999). «Viruses and apoptosis». Annual Review of Microbiology. 53: 577–628. doi:10.1146/annurev.micro.53.1.577. PMID 10547702.
- ^ Madigan MT, Martino JM (2006). Brock Biology of Microorganisms (11th ed.). Pearson. p. 136. ISBN 978-0-13-196893-6.
- ^ Ohta T, Tokishita SI, Mochizuki K, Kawase J, Sakahira M, Yamagata H (2006). «UV Sensitivity and Mutagenesis of the Extremely Thermophilic Eubacterium Thermus thermophilus HB27». Genes and Environment. 28 (2): 56–61. doi:10.3123/jemsge.28.56.
- ^ Tanaka T, Halicka HD, Huang X, Traganos F, Darzynkiewicz Z (September 2006). «Constitutive histone H2AX phosphorylation and ATM activation, the reporters of DNA damage by endogenous oxidants». Cell Cycle. 5 (17): 1940–45. doi:10.4161/cc.5.17.3191. PMC 3488278. PMID 16940754.
- ^ Braig M, Schmitt CA (March 2006). «Oncogene-induced senescence: putting the brakes on tumor development». Cancer Research. 66 (6): 2881–84. doi:10.1158/0008-5472.CAN-05-4006. PMID 16540631.
- ^ Lynch MD (February 2006). «How does cellular senescence prevent cancer?». DNA and Cell Biology. 25 (2): 69–78. doi:10.1089/dna.2006.25.69. PMID 16460230.
- ^ Campisi J, d’Adda di Fagagna F (September 2007). «Cellular senescence: when bad things happen to good cells». Nature Reviews. Molecular Cell Biology. 8 (9): 729–40. doi:10.1038/nrm2233. PMID 17667954. S2CID 15664931.
- ^ a b c Best BP (June 2009). «Nuclear DNA damage as a direct cause of aging» (PDF). Rejuvenation Research. 12 (3): 199–208. CiteSeerX 10.1.1.318.738. doi:10.1089/rej.2009.0847. PMID 19594328. Archived from the original (PDF) on 15 November 2017. Retrieved 29 September 2009.
- ^ Sancar A (June 2003). «Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors». Chemical Reviews. 103 (6): 2203–37. doi:10.1021/cr0204348. PMID 12797829.
- ^ Lucas-Lledó JI, Lynch M (May 2009). «Evolution of mutation rates: phylogenomic analysis of the photolyase/cryptochrome family». Molecular Biology and Evolution. 26 (5): 1143–53. doi:10.1093/molbev/msp029. PMC 2668831. PMID 19228922.
- ^ a b c Watson JD, Baker TA, Bell SP, Gann A, Levine M, Losick R (2004). Molecular Biology of the Gene (5th ed.). Pearson Benjamin Cummings; CSHL Press. Ch. 9, 10. OCLC 936762772.
- ^ Volkert MR (1988). «Adaptive response of Escherichia coli to alkylation damage». Environmental and Molecular Mutagenesis. 11 (2): 241–55. doi:10.1002/em.2850110210. PMID 3278898. S2CID 24722637.
- ^ a b c Willey J, Sherwood L, Woolverton C (2014). Prescott’s Microbiology. New York: McGraw Hill. p. 381. ISBN 978-0-07-3402-40-6.
- ^ Russell P (2018). i Genetics. Chennai: Pearson. p. 186. ISBN 978-93-325-7162-4.
- ^ a b c d Reardon JT, Sancar A (2006). «Purification and characterization of Escherichia coli and human nucleotide excision repair enzyme systems». Methods in Enzymology. 408: 189–213. doi:10.1016/S0076-6879(06)08012-8. ISBN 9780121828134. PMID 16793370.
- ^ Berg M, Tymoczko J, Stryer L (2012). Biochemistry 7th edition. New York: W.H. Freeman and Company. p. 840. ISBN 9781429229364.
- ^ a b Liang L, Deng L, Chen Y, Li GC, Shao C, Tischfield JA (September 2005). «Modulation of DNA end joining by nuclear proteins». The Journal of Biological Chemistry. 280 (36): 31442–49. doi:10.1074/jbc.M503776200. PMID 16012167.
- ^ Wilson TE, Grawunder U, Lieber MR (July 1997). «Yeast DNA ligase IV mediates non-homologous DNA end joining». Nature. 388 (6641): 495–98. Bibcode:1997Natur.388..495W. doi:10.1038/41365. PMID 9242411. S2CID 4422938.
- ^ Moore JK, Haber JE (May 1996). «Cell cycle and genetic requirements of two pathways of nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae». Molecular and Cellular Biology. 16 (5): 2164–73. doi:10.1128/mcb.16.5.2164. PMC 231204. PMID 8628283.
- ^ Boulton SJ, Jackson SP (September 1996). «Saccharomyces cerevisiae Ku70 potentiates illegitimate DNA double-strand break repair and serves as a barrier to error-prone DNA repair pathways». The EMBO Journal. 15 (18): 5093–103. doi:10.1002/j.1460-2075.1996.tb00890.x. PMC 452249. PMID 8890183.
- ^ Wilson TE, Lieber MR (August 1999). «Efficient processing of DNA ends during yeast nonhomologous end joining. Evidence for a DNA polymerase beta (Pol4)-dependent pathway». The Journal of Biological Chemistry. 274 (33): 23599–609. doi:10.1074/jbc.274.33.23599. PMID 10438542.
- ^ Budman J, Chu G (February 2005). «Processing of DNA for nonhomologous end-joining by cell-free extract». The EMBO Journal. 24 (4): 849–60. doi:10.1038/sj.emboj.7600563. PMC 549622. PMID 15692565.
- ^ Wang H, Perrault AR, Takeda Y, Qin W, Wang H, Iliakis G (September 2003). «Biochemical evidence for Ku-independent backup pathways of NHEJ». Nucleic Acids Research. 31 (18): 5377–88. doi:10.1093/nar/gkg728. PMC 203313. PMID 12954774.
- ^ Jung D, Alt FW (January 2004). «Unraveling V(D)J recombination; insights into gene regulation». Cell. 116 (2): 299–311. doi:10.1016/S0092-8674(04)00039-X. PMID 14744439. S2CID 16890458.
- ^ a b Truong LN, Li Y, Shi LZ, Hwang PY, He J, Wang H, et al. (May 2013). «Microhomology-mediated End Joining and Homologous Recombination share the initial end resection step to repair DNA double-strand breaks in mammalian cells». Proceedings of the National Academy of Sciences of the United States of America. 110 (19): 7720–25. Bibcode:2013PNAS..110.7720T. doi:10.1073/pnas.1213431110. PMC 3651503. PMID 23610439.
- ^ Sharma S, Javadekar SM, Pandey M, Srivastava M, Kumari R, Raghavan SC (March 2015). «Homology and enzymatic requirements of microhomology-dependent alternative end joining». Cell Death & Disease. 6 (3): e1697. doi:10.1038/cddis.2015.58. PMC 4385936. PMID 25789972.
- ^ Decottignies A (2013). «Alternative end-joining mechanisms: a historical perspective». Frontiers in Genetics. 4: 48. doi:10.3389/fgene.2013.00048. PMC 3613618. PMID 23565119.
- ^ Zahradka K, Slade D, Bailone A, Sommer S, Averbeck D, Petranovic M, et al. (October 2006). «Reassembly of shattered chromosomes in Deinococcus radiodurans». Nature. 443 (7111): 569–73. Bibcode:2006Natur.443..569Z. doi:10.1038/nature05160. PMID 17006450. S2CID 4412830.
- ^ Stenerlöw B, Karlsson KH, Cooper B, Rydberg B. «Measurement of prompt DNA double-strand breaks in mammalian cells without including heat-labile sites: results for cells deficient in nonhomologous end joining». Radiat Res. 2003 Apr;159(4):502–10. doi:10.1667/0033-7587(2003)159[0502:mopdds2.0.co;2] PMID 12643795.
- ^ Waters LS, Minesinger BK, Wiltrout ME, D’Souza S, Woodruff RV, Walker GC (March 2009). «Eukaryotic translesion polymerases and their roles and regulation in DNA damage tolerance». Microbiology and Molecular Biology Reviews. 73 (1): 134–54. doi:10.1128/MMBR.00034-08. PMC 2650891. PMID 19258535.
- ^ Colis LC, Raychaudhury P, Basu AK (August 2008). «Mutational specificity of gamma-radiation-induced guanine-thymine and thymine-guanine intrastrand cross-links in mammalian cells and translesion synthesis past the guanine-thymine lesion by human DNA polymerase eta». Biochemistry. 47 (31): 8070–79. doi:10.1021/bi800529f. PMC 2646719. PMID 18616294.
- ^ Raychaudhury P, Basu AK (March 2011). «Genetic requirement for mutagenesis of the G[8,5-Me]T cross-link in Escherichia coli: DNA polymerases IV and V compete for error-prone bypass». Biochemistry. 50 (12): 2330–38. doi:10.1021/bi102064z. PMC 3062377. PMID 21302943.
- ^ «Translesion Synthesis». Research.chem.psu.edu. Archived from the original on 10 March 2012. Retrieved 14 August 2012.
- ^ Wang Z (July 2001). «Translesion synthesis by the UmuC family of DNA polymerases». Mutation Research. 486 (2): 59–70. doi:10.1016/S0921-8777(01)00089-1. PMID 11425512.
- ^ a b c Friedberg EC, Walker GC, Siede W, Wood RD, Schultz RA, Ellenberger T. (2006). DNA Repair and Mutagenesis, part 3. ASM Press. 2nd ed.
- ^ Liu B, Yip RK, Zhou Z (November 2012). «Chromatin remodeling, DNA damage repair and aging». Current Genomics. 13 (7): 533–47. doi:10.2174/138920212803251373. PMC 3468886. PMID 23633913.
- ^ Halicka HD, Zhao H, Podhorecka M, Traganos F, Darzynkiewicz Z (July 2009). «Cytometric detection of chromatin relaxation, an early reporter of DNA damage response». Cell Cycle. 8 (14): 2233–37. doi:10.4161/cc.8.14.8984. PMC 3856216. PMID 19502789.
- ^ a b c Sellou H, Lebeaupin T, Chapuis C, Smith R, Hegele A, Singh HR, et al. (December 2016). «The poly(ADP-ribose)-dependent chromatin remodeler Alc1 induces local chromatin relaxation upon DNA damage». Molecular Biology of the Cell. 27 (24): 3791–99. doi:10.1091/mbc.E16-05-0269. PMC 5170603. PMID 27733626.
- ^ a b Van Meter M, Simon M, Tombline G, May A, Morello TD, Hubbard BP, et al. (September 2016). «JNK Phosphorylates SIRT6 to Stimulate DNA Double-Strand Break Repair in Response to Oxidative Stress by Recruiting PARP1 to DNA Breaks». Cell Reports. 16 (10): 2641–50. doi:10.1016/j.celrep.2016.08.006. PMC 5089070. PMID 27568560.
- ^ a b Haince JF, McDonald D, Rodrigue A, Déry U, Masson JY, Hendzel MJ, Poirier GG (January 2008). «PARP1-dependent kinetics of recruitment of MRE11 and NBS1 proteins to multiple DNA damage sites». The Journal of Biological Chemistry. 283 (2): 1197–208. doi:10.1074/jbc.M706734200. PMID 18025084.
- ^ a b c Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM (March 1998). «DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139». The Journal of Biological Chemistry. 273 (10): 5858–68. doi:10.1074/jbc.273.10.5858. PMID 9488723.
- ^ Mailand N, Bekker-Jensen S, Faustrup H, Melander F, Bartek J, Lukas C, Lukas J (November 2007). «RNF8 ubiquitylates histones at DNA double-strand breaks and promotes assembly of repair proteins». Cell. 131 (5): 887–900. doi:10.1016/j.cell.2007.09.040. PMID 18001824. S2CID 14232192.
- ^ Luijsterburg MS, Acs K, Ackermann L, Wiegant WW, Bekker-Jensen S, Larsen DH, et al. (May 2012). «A new non-catalytic role for ubiquitin ligase RNF8 in unfolding higher-order chromatin structure». The EMBO Journal. 31 (11): 2511–27. doi:10.1038/emboj.2012.104. PMC 3365417. PMID 22531782.
- ^ a b Luijsterburg MS, Goedhart J, Moser J, Kool H, Geverts B, Houtsmuller AB, et al. (August 2007). «Dynamic in vivo interaction of DDB2 E3 ubiquitin ligase with UV-damaged DNA is independent of damage-recognition protein XPC». Journal of Cell Science. 120 (Pt 15): 2706–16. doi:10.1242/jcs.008367. PMID 17635991.
- ^ a b Pines A, Vrouwe MG, Marteijn JA, Typas D, Luijsterburg MS, Cansoy M, et al. (October 2012). «PARP1 promotes nucleotide excision repair through DDB2 stabilization and recruitment of ALC1». The Journal of Cell Biology. 199 (2): 235–49. doi:10.1083/jcb.201112132. PMC 3471223. PMID 23045548.
- ^ Jazayeri A, Falck J, Lukas C, Bartek J, Smith GC, Lukas J, Jackson SP (January 2006). «ATM- and cell cycle-dependent regulation of ATR in response to DNA double-strand breaks». Nature Cell Biology. 8 (1): 37–45. doi:10.1038/ncb1337. PMID 16327781. S2CID 9797133.
- ^ Bakkenist CJ, Kastan MB (January 2003). «DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation». Nature. 421 (6922): 499–506. Bibcode:2003Natur.421..499B. doi:10.1038/nature01368. PMID 12556884. S2CID 4403303.
- ^ Wei Q, Li L, Chen D (2007). DNA Repair, Genetic Instability, and Cancer. World Scientific. ISBN 978-981-270-014-8.[page needed]
- ^ Schonthal AH (2004). Checkpoint Controls and Cancer. Humana Press. ISBN 978-1-58829-500-2.[page needed]
- ^ Gartel AL, Tyner AL (June 2002). «The role of the cyclin-dependent kinase inhibitor p21 in apoptosis». Molecular Cancer Therapeutics. 1 (8): 639–49. PMID 12479224.
- ^ Janion C (2001). «Some aspects of the SOS response system—a critical survey». Acta Biochimica Polonica. 48 (3): 599–610. doi:10.18388/abp.2001_3894. PMID 11833768.
- ^ a b Erill I, Campoy S, Barbé J (November 2007). «Aeons of distress: an evolutionary perspective on the bacterial SOS response». FEMS Microbiology Reviews. 31 (6): 637–56. doi:10.1111/j.1574-6976.2007.00082.x. PMID 17883408.
- ^ Schlacher K, Pham P, Cox MM, Goodman MF (February 2006). «Roles of DNA polymerase V and RecA protein in SOS damage-induced mutation». Chemical Reviews. 106 (2): 406–19. doi:10.1021/cr0404951. PMID 16464012.
- ^ Fry RC, Begley TJ, Samson LD (2004). «Genome-wide responses to DNA-damaging agents». Annual Review of Microbiology. 59: 357–77. doi:10.1146/annurev.micro.59.031805.133658. PMID 16153173.
- ^ Espejel S, Martín M, Klatt P, Martín-Caballero J, Flores JM, Blasco MA (May 2004). «Shorter telomeres, accelerated ageing and increased lymphoma in DNA-PKcs-deficient mice». EMBO Reports. 5 (5): 503–09. doi:10.1038/sj.embor.7400127. PMC 1299048. PMID 15105825.
- ^ de Boer J, Andressoo JO, de Wit J, Huijmans J, Beems RB, van Steeg H, et al. (May 2002). «Premature aging in mice deficient in DNA repair and transcription». Science. 296 (5571): 1276–79. Bibcode:2002Sci…296.1276D. doi:10.1126/science.1070174. PMID 11950998. S2CID 41930529.
- ^ Dollé ME, Busuttil RA, Garcia AM, Wijnhoven S, van Drunen E, Niedernhofer LJ, et al. (April 2006). «Increased genomic instability is not a prerequisite for shortened lifespan in DNA repair deficient mice». Mutation Research. 596 (1–2): 22–35. doi:10.1016/j.mrfmmm.2005.11.008. PMID 16472827.
- ^ Kobayashi Y, Narumi I, Satoh K, Funayama T, Kikuchi M, Kitayama S, Watanabe H (November 2004). «Radiation response mechanisms of the extremely radioresistant bacterium Deinococcus radiodurans». Uchu Seibutsu Kagaku. 18 (3): 134–35. PMID 15858357.
- ^ Spindler SR (September 2005). «Rapid and reversible induction of the longevity, anticancer and genomic effects of caloric restriction». Mechanisms of Ageing and Development. 126 (9): 960–66. doi:10.1016/j.mad.2005.03.016. PMID 15927235. S2CID 7067036.
- ^ Halicka HD, Zhao H, Li J, Lee YS, Hsieh TC, Wu JM, Darzynkiewicz Z (December 2012). «Potential anti-aging agents suppress the level of constitutive mTOR- and DNA damage- signaling». Aging. 4 (12): 952–65. doi:10.18632/aging.100521. PMC 3615161. PMID 23363784.
- ^ Tissenbaum HA, Guarente L (March 2001). «Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans». Nature. 410 (6825): 227–30. Bibcode:2001Natur.410..227T. doi:10.1038/35065638. PMID 11242085. S2CID 4356885.
- ^ Cohen HY, Miller C, Bitterman KJ, Wall NR, Hekking B, Kessler B, et al. (July 2004). «Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase». Science. 305 (5682): 390–92. Bibcode:2004Sci…305..390C. doi:10.1126/science.1099196. PMID 15205477. S2CID 33503081.
- ^ Cabelof DC, Yanamadala S, Raffoul JJ, Guo Z, Soofi A, Heydari AR (March 2003). «Caloric restriction promotes genomic stability by induction of base excision repair and reversal of its age-related decline». DNA Repair. 2 (3): 295–307. doi:10.1016/S1568-7864(02)00219-7. PMID 12547392.
- ^ Stuart JA, Karahalil B, Hogue BA, Souza-Pinto NC, Bohr VA (March 2004). «Mitochondrial and nuclear DNA base excision repair are affected differently by caloric restriction». FASEB Journal. 18 (3): 595–97. doi:10.1096/fj.03-0890fje. PMID 14734635. S2CID 43118901.
- ^ Walker DW, McColl G, Jenkins NL, Harris J, Lithgow GJ (May 2000). «Evolution of lifespan in C. elegans». Nature. 405 (6784): 296–97. doi:10.1038/35012693. PMID 10830948. S2CID 4402039.
- ^ Johnson G (28 December 2010). «Unearthing Prehistoric Tumors, and Debate». The New York Times.
If we lived long enough, sooner or later we all would get cancer.
- ^ Alberts B, Johnson A, Lewis J, et al. (2002). «The Preventable Causes of Cancer». Molecular biology of the cell (4th ed.). New York: Garland Science. ISBN 978-0-8153-4072-0.
A certain irreducible background incidence of cancer is to be expected regardless of circumstances: mutations can never be absolutely avoided, because they are an inescapable consequence of fundamental limitations on the accuracy of DNA replication, as discussed in Chapter 5. If a human could live long enough, it is inevitable that at least one of his or her cells would eventually accumulate a set of mutations sufficient for cancer to develop.
- ^ Friedenson B (August 2007). «The BRCA1/2 pathway prevents hematologic cancers in addition to breast and ovarian cancers». BMC Cancer. 7: 152. doi:10.1186/1471-2407-7-152. PMC 1959234. PMID 17683622.
- ^ Gavande NS, VanderVere-Carozza PS, Hinshaw HD, Jalal SI, Sears CR, Pawelczak KS, Turchi JJ (April 2016). «DNA repair targeted therapy: The past or future of cancer treatment?». Pharmacology & Therapeutics. 160: 65–83. doi:10.1016/j.pharmthera.2016.02.003. PMC 4811676. PMID 26896565.
- ^ Bryant HE, Schultz N, Thomas HD, Parker KM, Flower D, Lopez E, et al. (April 2005). «Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase». Nature. 434 (7035): 913–17. Bibcode:2005Natur.434..913B. doi:10.1038/nature03443. PMID 15829966. S2CID 4391043.
- ^ Goldstein M, Kastan MB (2015). «The DNA damage response: implications for tumor responses to radiation and chemotherapy». Annual Review of Medicine. 66: 129–43. doi:10.1146/annurev-med-081313-121208. PMID 25423595.
- ^ a b Jeggo PA, Pearl LH, Carr AM (January 2016). «DNA repair, genome stability and cancer: a historical perspective» (PDF). Nature Reviews. Cancer. 16 (1): 35–42. doi:10.1038/nrc.2015.4. PMID 26667849. S2CID 14941857.
- ^ Bartkova J, Horejsí Z, Koed K, Krämer A, Tort F, Zieger K, et al. (April 2005). «DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis». Nature. 434 (7035): 864–70. Bibcode:2005Natur.434..864B. doi:10.1038/nature03482. PMID 15829956. S2CID 4398393.
- ^ a b Bartkova J, Rezaei N, Liontos M, Karakaidos P, Kletsas D, Issaeva N, et al. (November 2006). «Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints». Nature. 444 (7119): 633–37. Bibcode:2006Natur.444..633B. doi:10.1038/nature05268. PMID 17136093. S2CID 4406956.
- ^ Gaillard H, García-Muse T, Aguilera A (May 2015). «Replication stress and cancer». Nature Reviews. Cancer. 15 (5): 276–89. doi:10.1038/nrc3916. hdl:10261/123721. PMID 25907220. S2CID 11342123.
- ^ Halazonetis TD, Gorgoulis VG, Bartek J (March 2008). «An oncogene-induced DNA damage model for cancer development». Science. 319 (5868): 1352–55. Bibcode:2008Sci…319.1352H. doi:10.1126/science.1140735. PMID 18323444. S2CID 16426080.
- ^ de Boer J, Hoeijmakers JH (March 2000). «Nucleotide excision repair and human syndromes» (PDF). Carcinogenesis. 21 (3): 453–60. doi:10.1093/carcin/21.3.453. PMID 10688865.
- ^ Broustas CG, Lieberman HB (February 2014). «DNA damage response genes and the development of cancer metastasis». Radiation Research. 181 (2): 111–30. Bibcode:2014RadR..181..111B. doi:10.1667/RR13515.1. PMC 4064942. PMID 24397478.
- ^ Zhang P, Wang J, Gao W, Yuan BZ, Rogers J, Reed E (May 2004). «CHK2 kinase expression is down-regulated due to promoter methylation in non-small cell lung cancer». Molecular Cancer. 3 (4): 14. doi:10.1186/1476-4598-3-14. PMC 419366. PMID 15125777.
- ^ Baylin SB, Ohm JE (February 2006). «Epigenetic gene silencing in cancer – a mechanism for early oncogenic pathway addiction?». Nature Reviews. Cancer. 6 (2): 107–16. doi:10.1038/nrc1799. PMID 16491070. S2CID 2514545.
- ^ Kanwal R, Gupta S (April 2012). «Epigenetic modifications in cancer». Clinical Genetics. 81 (4): 303–11. doi:10.1111/j.1399-0004.2011.01809.x. PMC 3590802. PMID 22082348.
- ^ Baldassarre G, Battista S, Belletti B, Thakur S, Pentimalli F, Trapasso F, et al. (April 2003). «Negative regulation of BRCA1 gene expression by HMGA1 proteins accounts for the reduced BRCA1 protein levels in sporadic breast carcinoma». Molecular and Cellular Biology. 23 (7): 2225–38. doi:10.1128/MCB.23.7.2225-2238.2003. PMC 150734. PMID 12640109.
- ^ Jacinto FV, Esteller M (July 2007). «Mutator pathways unleashed by epigenetic silencing in human cancer». Mutagenesis. 22 (4): 247–53. doi:10.1093/mutage/gem009. PMID 17412712.
- ^ Lahtz C, Pfeifer GP (February 2011). «Epigenetic changes of DNA repair genes in cancer». Journal of Molecular Cell Biology. 3 (1): 51–58. doi:10.1093/jmcb/mjq053. PMC 3030973. PMID 21278452. Epigenetic changes of DNA repair genes in cancer
- ^ Bernstein C, Nfonsam V, Prasad AR, Bernstein H (March 2013). «Epigenetic field defects in progression to cancer». World Journal of Gastrointestinal Oncology. 5 (3): 43–49. doi:10.4251/wjgo.v5.i3.43. PMC 3648662. PMID 23671730.
- ^ Narayanan L, Fritzell JA, Baker SM, Liskay RM, Glazer PM (April 1997). «Elevated levels of mutation in multiple tissues of mice deficient in the DNA mismatch repair gene Pms2». Proceedings of the National Academy of Sciences of the United States of America. 94 (7): 3122–27. Bibcode:1997PNAS…94.3122N. doi:10.1073/pnas.94.7.3122. PMC 20332. PMID 9096356.
- ^ Hegan DC, Narayanan L, Jirik FR, Edelmann W, Liskay RM, Glazer PM (December 2006). «Differing patterns of genetic instability in mice deficient in the mismatch repair genes Pms2, Mlh1, Msh2, Msh3 and Msh6». Carcinogenesis. 27 (12): 2402–08. doi:10.1093/carcin/bgl079. PMC 2612936. PMID 16728433.
- ^ Tutt AN, van Oostrom CT, Ross GM, van Steeg H, Ashworth A (March 2002). «Disruption of Brca2 increases the spontaneous mutation rate in vivo: synergism with ionizing radiation». EMBO Reports. 3 (3): 255–60. doi:10.1093/embo-reports/kvf037. PMC 1084010. PMID 11850397.
- ^ German J (March 1969). «Bloom’s syndrome. I. Genetical and clinical observations in the first twenty-seven patients». American Journal of Human Genetics. 21 (2): 196–227. PMC 1706430. PMID 5770175.
- ^ O’Hagan HM, Mohammad HP, Baylin SB (August 2008). «Double strand breaks can initiate gene silencing and SIRT1-dependent onset of DNA methylation in an exogenous promoter CpG island». PLOS Genetics. 4 (8): e1000155. doi:10.1371/journal.pgen.1000155. PMC 2491723. PMID 18704159.
- ^ Cuozzo C, Porcellini A, Angrisano T, Morano A, Lee B, Di Pardo A, et al. (July 2007). «DNA damage, homology-directed repair, and DNA methylation». PLOS Genetics. 3 (7): e110. doi:10.1371/journal.pgen.0030110. PMC 1913100. PMID 17616978.
- ^ Malkin D (April 2011). «Li-fraumeni syndrome». Genes & Cancer. 2 (4): 475–84. doi:10.1177/1947601911413466. PMC 3135649. PMID 21779515.
- ^ Fearon ER (November 1997). «Human cancer syndromes: clues to the origin and nature of cancer». Science. 278 (5340): 1043–50. Bibcode:1997Sci…278.1043F. doi:10.1126/science.278.5340.1043. PMID 9353177.
- ^ Halford S, Rowan A, Sawyer E, Talbot I, Tomlinson I (June 2005). «O(6)-methylguanine methyltransferase in colorectal cancers: detection of mutations, loss of expression, and weak association with G:C>A:T transitions». Gut. 54 (6): 797–802. doi:10.1136/gut.2004.059535. PMC 1774551. PMID 15888787.
- ^ Shen L, Kondo Y, Rosner GL, Xiao L, Hernandez NS, Vilaythong J, et al. (September 2005). «MGMT promoter methylation and field defect in sporadic colorectal cancer». Journal of the National Cancer Institute. 97 (18): 1330–38. doi:10.1093/jnci/dji275. PMID 16174854.
- ^ Psofaki V, Kalogera C, Tzambouras N, Stephanou D, Tsianos E, Seferiadis K, Kolios G (July 2010). «Promoter methylation status of hMLH1, MGMT, and CDKN2A/p16 in colorectal adenomas». World Journal of Gastroenterology. 16 (28): 3553–60. doi:10.3748/wjg.v16.i28.3553. PMC 2909555. PMID 20653064.
- ^ Lee KH, Lee JS, Nam JH, Choi C, Lee MC, Park CS, et al. (October 2011). «Promoter methylation status of hMLH1, hMSH2, and MGMT genes in colorectal cancer associated with adenoma-carcinoma sequence». Langenbeck’s Archives of Surgery. 396 (7): 1017–26. doi:10.1007/s00423-011-0812-9. PMID 21706233. S2CID 8069716.
- ^ Amatu A, Sartore-Bianchi A, Moutinho C, Belotti A, Bencardino K, Chirico G, et al. (April 2013). «Promoter CpG island hypermethylation of the DNA repair enzyme MGMT predicts clinical response to dacarbazine in a phase II study for metastatic colorectal cancer». Clinical Cancer Research. 19 (8): 2265–72. doi:10.1158/1078-0432.CCR-12-3518. PMID 23422094.
- ^ Mokarram P, Zamani M, Kavousipour S, Naghibalhossaini F, Irajie C, Moradi Sarabi M, Hosseini SV (May 2013). «Different patterns of DNA methylation of the two distinct O6-methylguanine-DNA methyltransferase (O6-MGMT) promoter regions in colorectal cancer». Molecular Biology Reports. 40 (5): 3851–57. doi:10.1007/s11033-012-2465-3. PMID 23271133. S2CID 18733871.
- ^ Truninger K, Menigatti M, Luz J, Russell A, Haider R, Gebbers JO, et al. (May 2005). «Immunohistochemical analysis reveals high frequency of PMS2 defects in colorectal cancer». Gastroenterology. 128 (5): 1160–71. doi:10.1053/j.gastro.2005.01.056. PMID 15887099.
- ^ Valeri N, Gasparini P, Fabbri M, Braconi C, Veronese A, Lovat F, et al. (April 2010). «Modulation of mismatch repair and genomic stability by miR-155». Proceedings of the National Academy of Sciences of the United States of America. 107 (15): 6982–87. Bibcode:2010PNAS..107.6982V. doi:10.1073/pnas.1002472107. PMC 2872463. PMID 20351277.
- ^ Facista A, Nguyen H, Lewis C, Prasad AR, Ramsey L, Zaitlin B, et al. (April 2012). «Deficient expression of DNA repair enzymes in early progression to sporadic colon cancer». Genome Integrity. 3 (1): 3. doi:10.1186/2041-9414-3-3. PMC 3351028. PMID 22494821.
- ^ Human DNA Repair Genes, 15 April 2014, MD Anderson Cancer Center, University of Texas
- ^ Jin B, Robertson KD (2013). «DNA methyltransferases, DNA damage repair, and cancer». Advances in Experimental Medicine and Biology. 754: 3–29. doi:10.1007/978-1-4419-9967-2_1. ISBN 978-1-4419-9966-5. PMC 3707278. PMID 22956494.
- ^ Krishnan K, Steptoe AL, Martin HC, Wani S, Nones K, Waddell N, et al. (February 2013). «MicroRNA-182-5p targets a network of genes involved in DNA repair». RNA. 19 (2): 230–42. doi:10.1261/rna.034926.112. PMC 3543090. PMID 23249749.
- ^ Chaisaingmongkol J, Popanda O, Warta R, Dyckhoff G, Herpel E, Geiselhart L, et al. (December 2012). «Epigenetic screen of human DNA repair genes identifies aberrant promoter methylation of NEIL1 in head and neck squamous cell carcinoma». Oncogene. 31 (49): 5108–16. doi:10.1038/onc.2011.660. PMID 22286769.
- ^ Singh P, Yang M, Dai H, Yu D, Huang Q, Tan W, et al. (November 2008). «Overexpression and hypomethylation of flap endonuclease 1 gene in breast and other cancers». Molecular Cancer Research. 6 (11): 1710–17. doi:10.1158/1541-7786.MCR-08-0269. PMC 2948671. PMID 19010819.
- ^ Lam JS, Seligson DB, Yu H, Li A, Eeva M, Pantuck AJ, et al. (August 2006). «Flap endonuclease 1 is overexpressed in prostate cancer and is associated with a high Gleason score». BJU International. 98 (2): 445–51. doi:10.1111/j.1464-410X.2006.06224.x. PMID 16879693. S2CID 22165252.
- ^ Kim JM, Sohn HY, Yoon SY, Oh JH, Yang JO, Kim JH, et al. (January 2005). «Identification of gastric cancer-related genes using a cDNA microarray containing novel expressed sequence tags expressed in gastric cancer cells». Clinical Cancer Research. 11 (2 Pt 1): 473–82. doi:10.1158/1078-0432.473.11.2. PMID 15701830.
- ^ Wang K, Xie C, Chen D (May 2014). «Flap endonuclease 1 is a promising candidate biomarker in gastric cancer and is involved in cell proliferation and apoptosis». International Journal of Molecular Medicine. 33 (5): 1268–74. doi:10.3892/ijmm.2014.1682. PMID 24590400.
- ^ Krause A, Combaret V, Iacono I, Lacroix B, Compagnon C, Bergeron C, et al. (July 2005). «Genome-wide analysis of gene expression in neuroblastomas detected by mass screening» (PDF). Cancer Letters. 225 (1): 111–20. doi:10.1016/j.canlet.2004.10.035. PMID 15922863. S2CID 44644467.
- ^ Iacobuzio-Donahue CA, Maitra A, Olsen M, Lowe AW, van Heek NT, Rosty C, et al. (April 2003). «Exploration of global gene expression patterns in pancreatic adenocarcinoma using cDNA microarrays». The American Journal of Pathology. 162 (4): 1151–62. doi:10.1016/S0002-9440(10)63911-9. PMC 1851213. PMID 12651607.
- ^ Sato M, Girard L, Sekine I, Sunaga N, Ramirez RD, Kamibayashi C, Minna JD (October 2003). «Increased expression and no mutation of the Flap endonuclease (FEN1) gene in human lung cancer». Oncogene. 22 (46): 7243–46. doi:10.1038/sj.onc.1206977. PMID 14562054.
- ^ Bi FF, Li D, Yang Q (2013). «Hypomethylation of ETS transcription factor binding sites and upregulation of PARP1 expression in endometrial cancer». BioMed Research International. 2013: 946268. doi:10.1155/2013/946268. PMC 3666359. PMID 23762867.
- ^ Bi FF, Li D, Yang Q (February 2013). «Promoter hypomethylation, especially around the E26 transformation-specific motif, and increased expression of poly (ADP-ribose) polymerase 1 in BRCA-mutated serous ovarian cancer». BMC Cancer. 13: 90. doi:10.1186/1471-2407-13-90. PMC 3599366. PMID 23442605.
- ^ Supek F, Lehner B (May 2015). «Differential DNA mismatch repair underlies mutation rate variation across the human genome». Nature. 521 (7550): 81–84. Bibcode:2015Natur.521…81S. doi:10.1038/nature14173. PMC 4425546. PMID 25707793.
- ^ a b Zheng CL, Wang NJ, Chung J, Moslehi H, Sanborn JZ, Hur JS, et al. (November 2014). «Transcription restores DNA repair to heterochromatin, determining regional mutation rates in cancer genomes». Cell Reports. 9 (4): 1228–34. doi:10.1016/j.celrep.2014.10.031. PMC 4254608. PMID 25456125.
- ^ Li F, Mao G, Tong D, Huang J, Gu L, Yang W, Li GM (April 2013). «The histone mark H3K36me3 regulates human DNA mismatch repair through its interaction with MutSα». Cell. 153 (3): 590–600. doi:10.1016/j.cell.2013.03.025. PMC 3641580. PMID 23622243.
- ^ Supek F, Lehner B (July 2017). «Clustered Mutation Signatures Reveal that Error-Prone DNA Repair Targets Mutations to Active Genes». Cell. 170 (3): 534–547.e23. doi:10.1016/j.cell.2017.07.003. hdl:10230/35343. PMID 28753428.
- ^ Polak P, Lawrence MS, Haugen E, Stoletzki N, Stojanov P, Thurman RE, et al. (January 2014). «Reduced local mutation density in regulatory DNA of cancer genomes is linked to DNA repair». Nature Biotechnology. 32 (1): 71–75. doi:10.1038/nbt.2778. PMC 4116484. PMID 24336318.
- ^ Swenberg JA, Lu K, Moeller BC, Gao L, Upton PB, Nakamura J, Starr TB (March 2011). «Endogenous versus exogenous DNA adducts: their role in carcinogenesis, epidemiology, and risk assessment». Toxicol Sci. 120 Suppl 1 (Suppl 1): S130–45. doi:10.1093/toxsci/kfq371. PMC 3043087. PMID 21163908.
- ^ a b Hamilton ML, Guo Z, Fuller CD, Van Remmen H, Ward WF, Austad SN, Troyer DA, Thompson I, Richardson A (May 2001). «A reliable assessment of 8-oxo-2-deoxyguanosine levels in nuclear and mitochondrial DNA using the sodium iodide method to isolate DNA». Nucleic Acids Res. 29 (10): 2117–26. doi:10.1093/nar/29.10.2117. PMC 55450. PMID 11353081.
- ^ Ming X, Matter B, Song M, Veliath E, Shanley R, Jones R, Tretyakova N (March 2014). «Mapping structurally defined guanine oxidation products along DNA duplexes: influence of local sequence context and endogenous cytosine methylation». J Am Chem Soc. 136 (11): 4223–35. doi:10.1021/ja411636j. PMC 3985951. PMID 24571128.
- ^ a b c Zhou X, Zhuang Z, Wang W, He L, Wu H, Cao Y, Pan F, Zhao J, Hu Z, Sekhar C, Guo Z (September 2016). «OGG1 is essential in oxidative stress-induced DNA demethylation». Cell Signal. 28 (9): 1163–1171. doi:10.1016/j.cellsig.2016.05.021. PMID 27251462.
- ^ Poetsch AR (2020). «The genomics of oxidative DNA damage, repair, and resulting mutagenesis». Comput Struct Biotechnol J. 18: 207–219. doi:10.1016/j.csbj.2019.12.013. PMC 6974700. PMID 31993111.
- ^ D’Augustin O, Huet S, Campalans A, Radicella JP (November 2020). «Lost in the Crowd: How Does Human 8-Oxoguanine DNA Glycosylase 1 (OGG1) Find 8-Oxoguanine in the Genome?». Int J Mol Sci. 21 (21): 8360. doi:10.3390/ijms21218360. PMC 7664663. PMID 33171795.
- ^ Lan L, Nakajima S, Oohata Y, Takao M, Okano S, Masutani M, Wilson SH, Yasui A (September 2004). «In situ analysis of repair processes for oxidative DNA damage in mammalian cells». Proc Natl Acad Sci U S A. 101 (38): 13738–43. Bibcode:2004PNAS..10113738L. doi:10.1073/pnas.0406048101. PMC 518826. PMID 15365186.
- ^ Maeder ML, Angstman JF, Richardson ME, Linder SJ, Cascio VM, Tsai SQ, Ho QH, Sander JD, Reyon D, Bernstein BE, Costello JF, Wilkinson MF, Joung JK (December 2013). «Targeted DNA demethylation and activation of endogenous genes using programmable TALE-TET1 fusion proteins». Nat. Biotechnol. 31 (12): 1137–42. doi:10.1038/nbt.2726. PMC 3858462. PMID 24108092.
- ^ Ding N, Bonham EM, Hannon BE, Amick TR, Baylin SB, O’Hagan HM (June 2016). «Mismatch repair proteins recruit DNA methyltransferase 1 to sites of oxidative DNA damage». J Mol Cell Biol. 8 (3): 244–54. doi:10.1093/jmcb/mjv050. PMC 4937888. PMID 26186941.
- ^ a b Jiang Z, Lai Y, Beaver JM, Tsegay PS, Zhao ML, Horton JK, Zamora M, Rein HL, Miralles F, Shaver M, Hutcheson JD, Agoulnik I, Wilson SH, Liu Y (January 2020). «Oxidative DNA Damage Modulates DNA Methylation Pattern in Human Breast Cancer 1 (BRCA1) Gene via the Crosstalk between DNA Polymerase β and a de novo DNA Methyltransferase». Cells. 9 (1): 225. doi:10.3390/cells9010225. PMC 7016758. PMID 31963223.
- ^ Mortusewicz O, Schermelleh L, Walter J, Cardoso MC, Leonhardt H (June 2005). «Recruitment of DNA methyltransferase I to DNA repair sites». Proc Natl Acad Sci U S A. 102 (25): 8905–9. Bibcode:2005PNAS..102.8905M. doi:10.1073/pnas.0501034102. PMC 1157029. PMID 15956212.
- ^ a b Cuozzo C, Porcellini A, Angrisano T, Morano A, Lee B, Di Pardo A, Messina S, Iuliano R, Fusco A, Santillo MR, Muller MT, Chiariotti L, Gottesman ME, Avvedimento EV (July 2007). «DNA damage, homology-directed repair, and DNA methylation». PLOS Genet. 3 (7): e110. doi:10.1371/journal.pgen.0030110. PMC 1913100. PMID 17616978.
- ^ a b O’Hagan HM, Mohammad HP, Baylin SB (August 2008). «Double strand breaks can initiate gene silencing and SIRT1-dependent onset of DNA methylation in an exogenous promoter CpG island». PLOS Genet. 4 (8): e1000155. doi:10.1371/journal.pgen.1000155. PMC 2491723. PMID 18704159.
- ^ Ha K, Lee GE, Palii SS, Brown KD, Takeda Y, Liu K, Bhalla KN, Robertson KD (January 2011). «Rapid and transient recruitment of DNMT1 to DNA double-strand breaks is mediated by its interaction with multiple components of the DNA damage response machinery». Hum Mol Genet. 20 (1): 126–40. doi:10.1093/hmg/ddq451. PMC 3000680. PMID 20940144.
- ^ Russo G, Landi R, Pezone A, Morano A, Zuchegna C, Romano A, Muller MT, Gottesman ME, Porcellini A, Avvedimento EV (September 2016). «DNA damage and Repair Modify DNA methylation and Chromatin Domain of the Targeted Locus: Mechanism of allele methylation polymorphism». Sci Rep. 6: 33222. Bibcode:2016NatSR…633222R. doi:10.1038/srep33222. PMC 5024116. PMID 27629060.
- ^ Farris MH, Texter PA, Mora AA, Wiles MV, Mac Garrigle EF, Klaus SA, Rosfjord K (December 2020). «Detection of CRISPR-mediated genome modifications through altered methylation patterns of CpG islands». BMC Genomics. 21 (1): 856. doi:10.1186/s12864-020-07233-2. PMC 7709351. PMID 33267773.
- ^ Allen B, Pezone A, Porcellini A, Muller MT, Masternak MM (June 2017). «Non-homologous end joining induced alterations in DNA methylation: A source of permanent epigenetic change». Oncotarget. 8 (25): 40359–40372. doi:10.18632/oncotarget.16122. PMC 5522286. PMID 28423717.
- ^ Cromie GA, Connelly JC, Leach DR (December 2001). «Recombination at double-strand breaks and DNA ends: conserved mechanisms from phage to humans». Molecular Cell. 8 (6): 1163–74. doi:10.1016/S1097-2765(01)00419-1. PMID 11779493.
- ^ O’Brien PJ (February 2006). «Catalytic promiscuity and the divergent evolution of DNA repair enzymes». Chemical Reviews. 106 (2): 720–52. doi:10.1021/cr040481v. PMID 16464022.
- ^ Maresca B, Schwartz JH (January 2006). «Sudden origins: a general mechanism of evolution based on stress protein concentration and rapid environmental change». The Anatomical Record Part B: The New Anatomist. 289 (1): 38–46. doi:10.1002/ar.b.20089. PMID 16437551.
- ^ «CRISPR gene-editing tool has scientists thrilled – but nervous» CBC news. Author Kelly Crowe. 30 November 2015.
External links[edit]
![]()
This audio file was created from a revision of this article dated 17 June 2005, and does not reflect subsequent edits.
 Media related to DNA repair at Wikimedia Commons
Media related to DNA repair at Wikimedia Commons- Roswell Park Cancer Institute DNA Repair Lectures
- A comprehensive list of Human DNA Repair Genes
- 3D structures of some DNA repair enzymes
- Human DNA repair diseases
- DNA repair special interest group
- DNA Repair Archived 12 February 2018 at the Wayback Machine
- DNA Damage and DNA Repair
- Segmental Progeria
- DNA-damage repair; the good, the bad, and the ugly
DNA damage resulting in multiple broken chromosomes
DNA repair is a collection of processes by which a cell identifies and corrects damage to the DNA molecules that encode its genome.[1] In human cells, both normal metabolic activities and environmental factors such as radiation can cause DNA damage, resulting in tens of thousands of individual molecular lesions per cell per day.[2] Many of these lesions cause structural damage to the DNA molecule and can alter or eliminate the cell’s ability to transcribe the gene that the affected DNA encodes. Other lesions induce potentially harmful mutations in the cell’s genome, which affect the survival of its daughter cells after it undergoes mitosis. As a consequence, the DNA repair process is constantly active as it responds to damage in the DNA structure. When normal repair processes fail, and when cellular apoptosis does not occur, irreparable DNA damage may occur, including double-strand breaks and DNA crosslinkages (interstrand crosslinks or ICLs).[3][4] This can eventually lead to malignant tumors, or cancer as per the two hit hypothesis.
The rate of DNA repair is dependent on many factors, including the cell type, the age of the cell, and the extracellular environment. A cell that has accumulated a large amount of DNA damage, or one that no longer effectively repairs damage incurred to its DNA, can enter one of three possible states:
- an irreversible state of dormancy, known as senescence
- cell suicide, also known as apoptosis or programmed cell death
- unregulated cell division, which can lead to the formation of a tumor that is cancerous
The DNA repair ability of a cell is vital to the integrity of its genome and thus to the normal functionality of that organism. Many genes that were initially shown to influence life span have turned out to be involved in DNA damage repair and protection.[5]
Paul Modrich talks about himself and his work in DNA repair.
The 2015 Nobel Prize in Chemistry was awarded to Tomas Lindahl, Paul Modrich, and Aziz Sancar for their work on the molecular mechanisms of DNA repair processes.[6][7]
DNA damage[edit]
DNA damage, due to environmental factors and normal metabolic processes inside the cell, occurs at a rate of 10,000 to 1,000,000 molecular lesions per cell per day.[2] While this constitutes only 0.000165% of the human genome’s approximately 6 billion bases, unrepaired lesions in critical genes (such as tumor suppressor genes) can impede a cell’s ability to carry out its function and appreciably increase the likelihood of tumor formation and contribute to tumour heterogeneity.
The vast majority of DNA damage affects the primary structure of the double helix; that is, the bases themselves are chemically modified. These modifications can in turn disrupt the molecules’ regular helical structure by introducing non-native chemical bonds or bulky adducts that do not fit in the standard double helix. Unlike proteins and RNA, DNA usually lacks tertiary structure and therefore damage or disturbance does not occur at that level. DNA is, however, supercoiled and wound around «packaging» proteins called histones (in eukaryotes), and both superstructures are vulnerable to the effects of DNA damage.
Sources[edit]
DNA damage can be subdivided into two main types:
- endogenous damage such as attack by reactive oxygen species produced from normal metabolic byproducts (spontaneous mutation), especially the process of oxidative deamination
- also includes replication errors
- exogenous damage caused by external agents such as
- ultraviolet [UV 200–400 nm] radiation from the sun or other artificial light sources
- other radiation frequencies, including x-rays and gamma rays
- hydrolysis or thermal disruption
- certain plant toxins
- human-made mutagenic chemicals, especially aromatic compounds that act as DNA intercalating agents
- viruses[8]
The replication of damaged DNA before cell division can lead to the incorporation of wrong bases opposite damaged ones. Daughter cells that inherit these wrong bases carry mutations from which the original DNA sequence is unrecoverable (except in the rare case of a back mutation, for example, through gene conversion).
Types[edit]
There are several types of damage to DNA due to endogenous cellular processes:
- oxidation of bases [e.g. 8-oxo-7,8-dihydroguanine (8-oxoG)] and generation of DNA strand interruptions from reactive oxygen species,
- alkylation of bases (usually methylation), such as formation of 7-methylguanosine, 1-methyladenine, 6-O-Methylguanine
- hydrolysis of bases, such as deamination, depurination, and depyrimidination.
- «bulky adduct formation» (e.g., benzo[a]pyrene diol epoxide-dG adduct, aristolactam I-dA adduct)
- mismatch of bases, due to errors in DNA replication, in which the wrong DNA base is stitched into place in a newly forming DNA strand, or a DNA base is skipped over or mistakenly inserted.
- Monoadduct damage cause by change in single nitrogenous base of DNA
- Diadduct damage
Damage caused by exogenous agents comes in many forms. Some examples are:
- UV-B light causes crosslinking between adjacent cytosine and thymine bases creating pyrimidine dimers. This is called direct DNA damage.
- UV-A light creates mostly free radicals. The damage caused by free radicals is called indirect DNA damage.
- Ionizing radiation such as that created by radioactive decay or in cosmic rays causes breaks in DNA strands. Intermediate-level ionizing radiation may induce irreparable DNA damage (leading to replicational and transcriptional errors needed for neoplasia or may trigger viral interactions) leading to pre-mature aging and cancer.
- Thermal disruption at elevated temperature increases the rate of depurination (loss of purine bases from the DNA backbone) and single-strand breaks. For example, hydrolytic depurination is seen in the thermophilic bacteria, which grow in hot springs at 40–80 °C.[9][10] The rate of depurination (300 purine residues per genome per generation) is too high in these species to be repaired by normal repair machinery, hence a possibility of an adaptive response cannot be ruled out.
- Industrial chemicals such as vinyl chloride and hydrogen peroxide, and environmental chemicals such as polycyclic aromatic hydrocarbons found in smoke, soot and tar create a huge diversity of DNA adducts- ethenobases, oxidized bases, alkylated phosphotriesters and crosslinking of DNA, just to name a few.
UV damage, alkylation/methylation, X-ray damage and oxidative damage are examples of induced damage. Spontaneous damage can include the loss of a base, deamination, sugar ring puckering and tautomeric shift. Constitutive (spontaneous) DNA damage caused by endogenous oxidants can be detected as a low level of histone H2AX phosphorylation in untreated cells.[11]
Nuclear versus mitochondrial[edit]
In human cells, and eukaryotic cells in general, DNA is found in two cellular locations – inside the nucleus and inside the mitochondria. Nuclear DNA (nDNA) exists as chromatin during non-replicative stages of the cell cycle and is condensed into aggregate structures known as chromosomes during cell division. In either state the DNA is highly compacted and wound up around bead-like proteins called histones. Whenever a cell needs to express the genetic information encoded in its nDNA the required chromosomal region is unravelled, genes located therein are expressed, and then the region is condensed back to its resting conformation. Mitochondrial DNA (mtDNA) is located inside mitochondria organelles, exists in multiple copies, and is also tightly associated with a number of proteins to form a complex known as the nucleoid. Inside mitochondria, reactive oxygen species (ROS), or free radicals, byproducts of the constant production of adenosine triphosphate (ATP) via oxidative phosphorylation, create a highly oxidative environment that is known to damage mtDNA. A critical enzyme in counteracting the toxicity of these species is superoxide dismutase, which is present in both the mitochondria and cytoplasm of eukaryotic cells.
Senescence and apoptosis[edit]
Senescence, an irreversible process in which the cell no longer divides, is a protective response to the shortening of the chromosome ends, called telomeres. The telomeres are long regions of repetitive noncoding DNA that cap chromosomes and undergo partial degradation each time a cell undergoes division (see Hayflick limit).[12] In contrast, quiescence is a reversible state of cellular dormancy that is unrelated to genome damage (see cell cycle). Senescence in cells may serve as a functional alternative to apoptosis in cases where the physical presence of a cell for spatial reasons is required by the organism,[13] which serves as a «last resort» mechanism to prevent a cell with damaged DNA from replicating inappropriately in the absence of pro-growth cellular signaling. Unregulated cell division can lead to the formation of a tumor (see cancer), which is potentially lethal to an organism. Therefore, the induction of senescence and apoptosis is considered to be part of a strategy of protection against cancer.[14]
Mutation[edit]
It is important to distinguish between DNA damage and mutation, the two major types of error in DNA. DNA damage and mutation are fundamentally different. Damage results in physical abnormalities in the DNA, such as single- and double-strand breaks, 8-hydroxydeoxyguanosine residues, and polycyclic aromatic hydrocarbon adducts. DNA damage can be recognized by enzymes, and thus can be correctly repaired if redundant information, such as the undamaged sequence in the complementary DNA strand or in a homologous chromosome, is available for copying. If a cell retains DNA damage, transcription of a gene can be prevented, and thus translation into a protein will also be blocked. Replication may also be blocked or the cell may die.
In contrast to DNA damage, a mutation is a change in the base sequence of the DNA. A mutation cannot be recognized by enzymes once the base change is present in both DNA strands, and thus a mutation cannot be repaired. At the cellular level, mutations can cause alterations in protein function and regulation. Mutations are replicated when the cell replicates. In a population of cells, mutant cells will increase or decrease in frequency according to the effects of the mutation on the ability of the cell to survive and reproduce.
Although distinctly different from each other, DNA damage and mutation are related because DNA damage often causes errors of DNA synthesis during replication or repair; these errors are a major source of mutation.
Given these properties of DNA damage and mutation, it can be seen that DNA damage is a special problem in non-dividing or slowly-dividing cells, where unrepaired damage will tend to accumulate over time. On the other hand, in rapidly dividing cells, unrepaired DNA damage that does not kill the cell by blocking replication will tend to cause replication errors and thus mutation. The great majority of mutations that are not neutral in their effect are deleterious to a cell’s survival. Thus, in a population of cells composing a tissue with replicating cells, mutant cells will tend to be lost. However, infrequent mutations that provide a survival advantage will tend to clonally expand at the expense of neighboring cells in the tissue. This advantage to the cell is disadvantageous to the whole organism because such mutant cells can give rise to cancer. Thus, DNA damage in frequently dividing cells, because it gives rise to mutations, is a prominent cause of cancer. In contrast, DNA damage in infrequently-dividing cells is likely a prominent cause of aging.[15]
Mechanisms[edit]
Cells cannot function if DNA damage corrupts the integrity and accessibility of essential information in the genome (but cells remain superficially functional when non-essential genes are missing or damaged). Depending on the type of damage inflicted on the DNA’s double helical structure, a variety of repair strategies have evolved to restore lost information. If possible, cells use the unmodified complementary strand of the DNA or the sister chromatid as a template to recover the original information. Without access to a template, cells use an error-prone recovery mechanism known as translesion synthesis as a last resort.
Damage to DNA alters the spatial configuration of the helix, and such alterations can be detected by the cell. Once damage is localized, specific DNA repair molecules bind at or near the site of damage, inducing other molecules to bind and form a complex that enables the actual repair to take place.
Direct reversal[edit]
Cells are known to eliminate three types of damage to their DNA by chemically reversing it. These mechanisms do not require a template, since the types of damage they counteract can occur in only one of the four bases. Such direct reversal mechanisms are specific to the type of damage incurred and do not involve breakage of the phosphodiester backbone. The formation of pyrimidine dimers upon irradiation with UV light results in an abnormal covalent bond between adjacent pyrimidine bases. The photoreactivation process directly reverses this damage by the action of the enzyme photolyase, whose activation is obligately dependent on energy absorbed from blue/UV light (300–500 nm wavelength) to promote catalysis.[16] Photolyase, an old enzyme present in bacteria, fungi, and most animals no longer functions in humans,[17] who instead use nucleotide excision repair to repair damage from UV irradiation. Another type of damage, methylation of guanine bases, is directly reversed by the enzyme methyl guanine methyl transferase (MGMT), the bacterial equivalent of which is called ogt. This is an expensive process because each MGMT molecule can be used only once; that is, the reaction is stoichiometric rather than catalytic.[18] A generalized response to methylating agents in bacteria is known as the adaptive response and confers a level of resistance to alkylating agents upon sustained exposure by upregulation of alkylation repair enzymes.[19] The third type of DNA damage reversed by cells is certain methylation of the bases cytosine and adenine.
Single-strand damage[edit]
Structure of the base-excision repair enzyme uracil-DNA glycosylase excising a hydrolytically-produced uracil residue from DNA. The uracil residue is shown in yellow.
When only one of the two strands of a double helix has a defect, the other strand can be used as a template to guide the correction of the damaged strand. In order to repair damage to one of the two paired molecules of DNA, there exist a number of excision repair mechanisms that remove the damaged nucleotide and replace it with an undamaged nucleotide complementary to that found in the undamaged DNA strand.[18]
- Base excision repair (BER): damaged single bases or nucleotides are most commonly repaired by removing the base or the nucleotide involved and then inserting the correct base or nucleotide. In base excision repair, a glycosylase[20] enzyme removes the damaged base from the DNA by cleaving the bond between the base and the deoxyribose. These enzymes remove a single base to create an apurinic or apyrimidinic site (AP site).[20] Enzymes called AP endonucleases nick the damaged DNA backbone at the AP site. DNA polymerase then removes the damaged region using its 5’ to 3’ exonuclease activity and correctly synthesizes the new strand using the complementary strand as a template.[20] The gap is then sealed by enzyme DNA ligase.[21]
- Nucleotide excision repair (NER): bulky, helix-distorting damage, such as pyrimidine dimerization caused by UV light is usually repaired by a three-step process. First the damage is recognized, then 12-24 nucleotide-long strands of DNA are removed both upstream and downstream of the damage site by endonucleases, and the removed DNA region is then resynthesized.[22] NER is a highly evolutionarily conserved repair mechanism and is used in nearly all eukaryotic and prokaryotic cells.[22] In prokaryotes, NER is mediated by Uvr proteins.[22] In eukaryotes, many more proteins are involved, although the general strategy is the same.[22]
- Mismatch repair systems are present in essentially all cells to correct errors that are not corrected by proofreading. These systems consist of at least two proteins. One detects the mismatch, and the other recruits an endonuclease that cleaves the newly synthesized DNA strand close to the region of damage. In E. coli , the proteins involved are the Mut class proteins: MutS, MutL, and MutH. In most Eukaryotes, the analog for MutS is MSH and the analog for MutL is MLH. MutH is only present in bacteria. This is followed by removal of damaged region by an exonuclease, resynthesis by DNA polymerase, and nick sealing by DNA ligase.[23]
Double-strand breaks[edit]
The main double-strand break repair pathways
Double-strand breaks, in which both strands in the double helix are severed, are particularly hazardous to the cell because they can lead to genome rearrangements. In fact, when a double-strand break is accompanied by a cross-linkage joining the two strands at the same point, neither strand can be used as a template for the repair mechanisms, so that the cell will not be able to complete mitosis when it next divides, and will either die or, in rare cases, undergo a mutation.[3][4] Three mechanisms exist to repair double-strand breaks (DSBs): non-homologous end joining (NHEJ), microhomology-mediated end joining (MMEJ), and homologous recombination (HR):[18][24]
DNA ligase, shown above repairing chromosomal damage, is an enzyme that joins broken nucleotides together by catalyzing the formation of an internucleotide ester bond between the phosphate backbone and the deoxyribose nucleotides.
- In NHEJ, DNA Ligase IV, a specialized DNA ligase that forms a complex with the cofactor XRCC4, directly joins the two ends.[25] To guide accurate repair, NHEJ relies on short homologous sequences called microhomologies present on the single-stranded tails of the DNA ends to be joined. If these overhangs are compatible, repair is usually accurate.[26][27][28][29] NHEJ can also introduce mutations during repair. Loss of damaged nucleotides at the break site can lead to deletions, and joining of nonmatching termini forms insertions or translocations. NHEJ is especially important before the cell has replicated its DNA, since there is no template available for repair by homologous recombination. There are «backup» NHEJ pathways in higher eukaryotes.[30] Besides its role as a genome caretaker, NHEJ is required for joining hairpin-capped double-strand breaks induced during V(D)J recombination, the process that generates diversity in B-cell and T-cell receptors in the vertebrate immune system.[31]
- MMEJ starts with short-range end resection by MRE11 nuclease on either side of a double-strand break to reveal microhomology regions.[32] In further steps,[33] Poly (ADP-ribose) polymerase 1 (PARP1) is required and may be an early step in MMEJ. There is pairing of microhomology regions followed by recruitment of flap structure-specific endonuclease 1 (FEN1) to remove overhanging flaps. This is followed by recruitment of XRCC1–LIG3 to the site for ligating the DNA ends, leading to an intact DNA. MMEJ is always accompanied by a deletion, so that MMEJ is a mutagenic pathway for DNA repair.[34]
- HR requires the presence of an identical or nearly identical sequence to be used as a template for repair of the break. The enzymatic machinery responsible for this repair process is nearly identical to the machinery responsible for chromosomal crossover during meiosis. This pathway allows a damaged chromosome to be repaired using a sister chromatid (available in G2 after DNA replication) or a homologous chromosome as a template. DSBs caused by the replication machinery attempting to synthesize across a single-strand break or unrepaired lesion cause collapse of the replication fork and are typically repaired by recombination.
In an in vitro system, MMEJ occurred in mammalian cells at the levels of 10–20% of HR when both HR and NHEJ mechanisms were also available.[32]
The extremophile Deinococcus radiodurans has a remarkable ability to survive DNA damage from ionizing radiation and other sources. At least two copies of the genome, with random DNA breaks, can form DNA fragments through annealing. Partially overlapping fragments are then used for synthesis of homologous regions through a moving D-loop that can continue extension until complementary partner strands are found. In the final step, there is crossover by means of RecA-dependent homologous recombination.[35]
Topoisomerases introduce both single- and double-strand breaks in the course of changing the DNA’s state of supercoiling, which is especially common in regions near an open replication fork. Such breaks are not considered DNA damage because they are a natural intermediate in the topoisomerase biochemical mechanism and are immediately repaired by the enzymes that created them.
Another type of DNA double-strand breaks originates from the DNA heat-sensitive or heat-labile sites. These DNA sites are not initial DSBs. However, they convert to DSB after treating with elevated temperature. Ionizing irradiation can induces a highly complex form of DNA damage as clustered damage. It consists of different types of DNA lesions in various locations of the DNA helix. Some of these closely located lesions can probably convert to DSB by exposure to high temperatures. But the exact nature of these lesions and their interactions is not yet known[36]
Translesion synthesis[edit]
Translesion synthesis (TLS) is a DNA damage tolerance process that allows the DNA replication machinery to replicate past DNA lesions such as thymine dimers or AP sites.[37] It involves switching out regular DNA polymerases for specialized translesion polymerases (i.e. DNA polymerase IV or V, from the Y Polymerase family), often with larger active sites that can facilitate the insertion of bases opposite damaged nucleotides. The polymerase switching is thought to be mediated by, among other factors, the post-translational modification of the replication processivity factor PCNA. Translesion synthesis polymerases often have low fidelity (high propensity to insert wrong bases) on undamaged templates relative to regular polymerases. However, many are extremely efficient at inserting correct bases opposite specific types of damage. For example, Pol η mediates error-free bypass of lesions induced by UV irradiation, whereas Pol ι introduces mutations at these sites. Pol η is known to add the first adenine across the T^T photodimer using Watson-Crick base pairing and the second adenine will be added in its syn conformation using Hoogsteen base pairing. From a cellular perspective, risking the introduction of point mutations during translesion synthesis may be preferable to resorting to more drastic mechanisms of DNA repair, which may cause gross chromosomal aberrations or cell death. In short, the process involves specialized polymerases either bypassing or repairing lesions at locations of stalled DNA replication. For example, Human DNA polymerase eta can bypass complex DNA lesions like guanine-thymine intra-strand crosslink, G[8,5-Me]T, although it can cause targeted and semi-targeted mutations.[38] Paromita Raychaudhury and Ashis Basu[39] studied the toxicity and mutagenesis of the same lesion in Escherichia coli by replicating a G[8,5-Me]T-modified plasmid in E. coli with specific DNA polymerase knockouts. Viability was very low in a strain lacking pol II, pol IV, and pol V, the three SOS-inducible DNA polymerases, indicating that translesion synthesis is conducted primarily by these specialized DNA polymerases.
A bypass platform is provided to these polymerases by Proliferating cell nuclear antigen (PCNA). Under normal circumstances, PCNA bound to polymerases replicates the DNA. At a site of lesion, PCNA is ubiquitinated, or modified, by the RAD6/RAD18 proteins to provide a platform for the specialized polymerases to bypass the lesion and resume DNA replication.[40][41] After translesion synthesis, extension is required. This extension can be carried out by a replicative polymerase if the TLS is error-free, as in the case of Pol η, yet if TLS results in a mismatch, a specialized polymerase is needed to extend it; Pol ζ. Pol ζ is unique in that it can extend terminal mismatches, whereas more processive polymerases cannot. So when a lesion is encountered, the replication fork will stall, PCNA will switch from a processive polymerase to a TLS polymerase such as Pol ι to fix the lesion, then PCNA may switch to Pol ζ to extend the mismatch, and last PCNA will switch to the processive polymerase to continue replication.
Global response to DNA damage[edit]
Cells exposed to ionizing radiation, ultraviolet light or chemicals are prone to acquire multiple sites of bulky DNA lesions and double-strand breaks. Moreover, DNA damaging agents can damage other biomolecules such as proteins, carbohydrates, lipids, and RNA. The accumulation of damage, to be specific, double-strand breaks or adducts stalling the replication forks, are among known stimulation signals for a global response to DNA damage.[42] The global response to damage is an act directed toward the cells’ own preservation and triggers multiple pathways of macromolecular repair, lesion bypass, tolerance, or apoptosis. The common features of global response are induction of multiple genes, cell cycle arrest, and inhibition of cell division.
Initial steps[edit]
The packaging of eukaryotic DNA into chromatin presents a barrier to all DNA-based processes that require recruitment of enzymes to their sites of action. To allow DNA repair, the chromatin must be remodeled. In eukaryotes, ATP dependent chromatin remodeling complexes and histone-modifying enzymes are two predominant factors employed to accomplish this remodeling process.[43]
Chromatin relaxation occurs rapidly at the site of a DNA damage.[44][45] In one of the earliest steps, the stress-activated protein kinase, c-Jun N-terminal kinase (JNK), phosphorylates SIRT6 on serine 10 in response to double-strand breaks or other DNA damage.[46] This post-translational modification facilitates the mobilization of SIRT6 to DNA damage sites, and is required for efficient recruitment of poly (ADP-ribose) polymerase 1 (PARP1) to DNA break sites and for efficient repair of DSBs.[46] PARP1 protein starts to appear at DNA damage sites in less than a second, with half maximum accumulation within 1.6 seconds after the damage occurs.[47] PARP1 synthesizes polymeric adenosine diphosphate ribose (poly (ADP-ribose) or PAR) chains on itself. Next the chromatin remodeler ALC1 quickly attaches to the product of PARP1 action, a poly-ADP ribose chain, and ALC1 completes arrival at the DNA damage within 10 seconds of the occurrence of the damage.[45] About half of the maximum chromatin relaxation, presumably due to action of ALC1, occurs by 10 seconds.[45] This then allows recruitment of the DNA repair enzyme MRE11, to initiate DNA repair, within 13 seconds.[47]
γH2AX, the phosphorylated form of H2AX is also involved in the early steps leading to chromatin decondensation after DNA double-strand breaks. The histone variant H2AX constitutes about 10% of the H2A histones in human chromatin.[48] γH2AX (H2AX phosphorylated on serine 139) can be detected as soon as 20 seconds after irradiation of cells (with DNA double-strand break formation), and half maximum accumulation of γH2AX occurs in one minute.[48] The extent of chromatin with phosphorylated γH2AX is about two million base pairs at the site of a DNA double-strand break.[48] γH2AX does not, itself, cause chromatin decondensation, but within 30 seconds of irradiation, RNF8 protein can be detected in association with γH2AX.[49] RNF8 mediates extensive chromatin decondensation, through its subsequent interaction with CHD4,[50] a component of the nucleosome remodeling and deacetylase complex NuRD.
DDB2 occurs in a heterodimeric complex with DDB1. This complex further complexes with the ubiquitin ligase protein CUL4A[51] and with PARP1.[52] This larger complex rapidly associates with UV-induced damage within chromatin, with half-maximum association completed in 40 seconds.[51] The PARP1 protein, attached to both DDB1 and DDB2, then PARylates (creates a poly-ADP ribose chain) on DDB2 that attracts the DNA remodeling protein ALC1.[52] Action of ALC1 relaxes the chromatin at the site of UV damage to DNA. This relaxation allows other proteins in the nucleotide excision repair pathway to enter the chromatin and repair UV-induced cyclobutane pyrimidine dimer damages.
After rapid chromatin remodeling, cell cycle checkpoints are activated to allow DNA repair to occur before the cell cycle progresses. First, two kinases, ATM and ATR are activated within 5 or 6 minutes after DNA is damaged. This is followed by phosphorylation of the cell cycle checkpoint protein Chk1, initiating its function, about 10 minutes after DNA is damaged.[53]
DNA damage checkpoints[edit]
After DNA damage, cell cycle checkpoints are activated. Checkpoint activation pauses the cell cycle and gives the cell time to repair the damage before continuing to divide. DNA damage checkpoints occur at the G1/S and G2/M boundaries. An intra-S checkpoint also exists. Checkpoint activation is controlled by two master kinases, ATM and ATR. ATM responds to DNA double-strand breaks and disruptions in chromatin structure,[54] whereas ATR primarily responds to stalled replication forks. These kinases phosphorylate downstream targets in a signal transduction cascade, eventually leading to cell cycle arrest. A class of checkpoint mediator proteins including BRCA1, MDC1, and 53BP1 has also been identified.[55] These proteins seem to be required for transmitting the checkpoint activation signal to downstream proteins.
DNA damage checkpoint is a signal transduction pathway that blocks cell cycle progression in G1, G2 and metaphase and slows down the rate of S phase progression when DNA is damaged. It leads to a pause in cell cycle allowing the cell time to repair the damage before continuing to divide.
Checkpoint Proteins can be separated into four groups: phosphatidylinositol 3-kinase (PI3K)-like protein kinase, proliferating cell nuclear antigen (PCNA)-like group, two serine/threonine(S/T) kinases and their adaptors. Central to all DNA damage induced checkpoints responses is a pair of large protein kinases belonging to the first group of PI3K-like protein kinases-the ATM (Ataxia telangiectasia mutated) and ATR (Ataxia- and Rad-related) kinases, whose sequence and functions have been well conserved in evolution. All DNA damage response requires either ATM or ATR because they have the ability to bind to the chromosomes at the site of DNA damage, together with accessory proteins that are platforms on which DNA damage response components and DNA repair complexes can be assembled.
An important downstream target of ATM and ATR is p53, as it is required for inducing apoptosis following DNA damage.[56] The cyclin-dependent kinase inhibitor p21 is induced by both p53-dependent and p53-independent mechanisms and can arrest the cell cycle at the G1/S and G2/M checkpoints by deactivating cyclin/cyclin-dependent kinase complexes.[57]
The prokaryotic SOS response[edit]
The SOS response is the changes in gene expression in Escherichia coli and other bacteria in response to extensive DNA damage. The prokaryotic SOS system is regulated by two key proteins: LexA and RecA. The LexA homodimer is a transcriptional repressor that binds to operator sequences commonly referred to as SOS boxes. In Escherichia coli it is known that LexA regulates transcription of approximately 48 genes including the lexA and recA genes.[58] The SOS response is known to be widespread in the Bacteria domain, but it is mostly absent in some bacterial phyla, like the Spirochetes.[59]
The most common cellular signals activating the SOS response are regions of single-stranded DNA (ssDNA), arising from stalled replication forks or double-strand breaks, which are processed by DNA helicase to separate the two DNA strands.[42] In the initiation step, RecA protein binds to ssDNA in an ATP hydrolysis driven reaction creating RecA–ssDNA filaments. RecA–ssDNA filaments activate LexA autoprotease activity, which ultimately leads to cleavage of LexA dimer and subsequent LexA degradation. The loss of LexA repressor induces transcription of the SOS genes and allows for further signal induction, inhibition of cell division and an increase in levels of proteins responsible for damage processing.
In Escherichia coli, SOS boxes are 20-nucleotide long sequences near promoters with palindromic structure and a high degree of sequence conservation. In other classes and phyla, the sequence of SOS boxes varies considerably, with different length and composition, but it is always highly conserved and one of the strongest short signals in the genome.[59] The high information content of SOS boxes permits differential binding of LexA to different promoters and allows for timing of the SOS response. The lesion repair genes are induced at the beginning of SOS response. The error-prone translesion polymerases, for example, UmuCD’2 (also called DNA polymerase V), are induced later on as a last resort.[60] Once the DNA damage is repaired or bypassed using polymerases or through recombination, the amount of single-stranded DNA in cells is decreased, lowering the amounts of RecA filaments decreases cleavage activity of LexA homodimer, which then binds to the SOS boxes near promoters and restores normal gene expression.
Eukaryotic transcriptional responses to DNA damage[edit]
Eukaryotic cells exposed to DNA damaging agents also activate important defensive pathways by inducing multiple proteins involved in DNA repair, cell cycle checkpoint control, protein trafficking and degradation. Such genome wide transcriptional response is very complex and tightly regulated, thus allowing coordinated global response to damage. Exposure of yeast Saccharomyces cerevisiae to DNA damaging agents results in overlapping but distinct transcriptional profiles. Similarities to environmental shock response indicates that a general global stress response pathway exist at the level of transcriptional activation. In contrast, different human cell types respond to damage differently indicating an absence of a common global response. The probable explanation for this difference between yeast and human cells may be in the heterogeneity of mammalian cells. In an animal different types of cells are distributed among different organs that have evolved different sensitivities to DNA damage.[61]
In general global response to DNA damage involves expression of multiple genes responsible for postreplication repair, homologous recombination, nucleotide excision repair, DNA damage checkpoint, global transcriptional activation, genes controlling mRNA decay, and many others. A large amount of damage to a cell leaves it with an important decision: undergo apoptosis and die, or survive at the cost of living with a modified genome. An increase in tolerance to damage can lead to an increased rate of survival that will allow a greater accumulation of mutations. Yeast Rev1 and human polymerase η are members of Y family translesion DNA polymerases present during global response to DNA damage and are responsible for enhanced mutagenesis during a global response to DNA damage in eukaryotes.[42]
Aging[edit]
Pathological effects of poor DNA repair[edit]
DNA repair rate is an important determinant of cell pathology.
Experimental animals with genetic deficiencies in DNA repair often show decreased life span and increased cancer incidence.[15] For example, mice deficient in the dominant NHEJ pathway and in telomere maintenance mechanisms get lymphoma and infections more often, and, as a consequence, have shorter lifespans than wild-type mice.[62] In similar manner, mice deficient in a key repair and transcription protein that unwinds DNA helices have premature onset of aging-related diseases and consequent shortening of lifespan.[63] However, not every DNA repair deficiency creates exactly the predicted effects; mice deficient in the NER pathway exhibited shortened life span without correspondingly higher rates of mutation.[64]
If the rate of DNA damage exceeds the capacity of the cell to repair it, the accumulation of errors can overwhelm the cell and result in early senescence, apoptosis, or cancer. Inherited diseases associated with faulty DNA repair functioning result in premature aging,[15] increased sensitivity to carcinogens and correspondingly increased cancer risk (see below). On the other hand, organisms with enhanced DNA repair systems, such as Deinococcus radiodurans, the most radiation-resistant known organism, exhibit remarkable resistance to the double-strand break-inducing effects of radioactivity, likely due to enhanced efficiency of DNA repair and especially NHEJ.[65]
Longevity and caloric restriction[edit]
Most life span influencing genes affect the rate of DNA damage.
A number of individual genes have been identified as influencing variations in life span within a population of organisms. The effects of these genes is strongly dependent on the environment, in particular, on the organism’s diet. Caloric restriction reproducibly results in extended lifespan in a variety of organisms, likely via nutrient sensing pathways and decreased metabolic rate. The molecular mechanisms by which such restriction results in lengthened lifespan are as yet unclear (see[66] for some discussion); however, the behavior of many genes known to be involved in DNA repair is altered under conditions of caloric restriction. Several agents reported to have anti-aging properties have been shown to attenuate constitutive level of mTOR signaling, an evidence of reduction of metabolic activity, and concurrently to reduce constitutive level of DNA damage induced by endogenously generated reactive oxygen species.[67]
For example, increasing the gene dosage of the gene SIR-2, which regulates DNA packaging in the nematode worm Caenorhabditis elegans, can significantly extend lifespan.[68] The mammalian homolog of SIR-2 is known to induce downstream DNA repair factors involved in NHEJ, an activity that is especially promoted under conditions of caloric restriction.[69] Caloric restriction has been closely linked to the rate of base excision repair in the nuclear DNA of rodents,[70] although similar effects have not been observed in mitochondrial DNA.[71]
The C. elegans gene AGE-1, an upstream effector of DNA repair pathways, confers dramatically extended life span under free-feeding conditions but leads to a decrease in reproductive fitness under conditions of caloric restriction.[72] This observation supports the pleiotropy theory of the biological origins of aging, which suggests that genes conferring a large survival advantage early in life will be selected for even if they carry a corresponding disadvantage late in life.
Medicine and DNA repair modulation[edit]
Hereditary DNA repair disorders[edit]
Defects in the NER mechanism are responsible for several genetic disorders, including:
- Xeroderma pigmentosum: hypersensitivity to sunlight/UV, resulting in increased skin cancer incidence and premature aging
- Cockayne syndrome: hypersensitivity to UV and chemical agents
- Trichothiodystrophy: sensitive skin, brittle hair and nails
Mental retardation often accompanies the latter two disorders, suggesting increased vulnerability of developmental neurons.
Other DNA repair disorders include:
- Werner’s syndrome: premature aging and retarded growth
- Bloom’s syndrome: sunlight hypersensitivity, high incidence of malignancies (especially leukemias).
- Ataxia telangiectasia: sensitivity to ionizing radiation and some chemical agents
All of the above diseases are often called «segmental progerias» («accelerated aging diseases») because those affected appear elderly and experience aging-related diseases at an abnormally young age, while not manifesting all the symptoms of old age.
Other diseases associated with reduced DNA repair function include Fanconi anemia, hereditary breast cancer and hereditary colon cancer.
Cancer[edit]
Because of inherent limitations in the DNA repair mechanisms, if humans lived long enough, they would all eventually develop cancer.[73][74] There are at least 34 Inherited human DNA repair gene mutations that increase cancer risk. Many of these mutations cause DNA repair to be less effective than normal. In particular, Hereditary nonpolyposis colorectal cancer (HNPCC) is strongly associated with specific mutations in the DNA mismatch repair pathway. BRCA1 and BRCA2, two important genes whose mutations confer a hugely increased risk of breast cancer on carriers,[75] are both associated with a large number of DNA repair pathways, especially NHEJ and homologous recombination.
Cancer therapy procedures such as chemotherapy and radiotherapy work by overwhelming the capacity of the cell to repair DNA damage, resulting in cell death. Cells that are most rapidly dividing – most typically cancer cells – are preferentially affected. The side-effect is that other non-cancerous but rapidly dividing cells such as progenitor cells in the gut, skin, and hematopoietic system are also affected. Modern cancer treatments attempt to localize the DNA damage to cells and tissues only associated with cancer, either by physical means (concentrating the therapeutic agent in the region of the tumor) or by biochemical means (exploiting a feature unique to cancer cells in the body). In the context of therapies targeting DNA damage response genes, the latter approach has been termed ‘synthetic lethality’.[76]
Perhaps the most well-known of these ‘synthetic lethality’ drugs is the poly(ADP-ribose) polymerase 1 (PARP1) inhibitor olaparib, which was approved by the Food and Drug Administration in 2015 for the treatment in women of BRCA-defective ovarian cancer. Tumor cells with partial loss of DNA damage response (specifically, homologous recombination repair) are dependent on another mechanism – single-strand break repair – which is a mechanism consisting, in part, of the PARP1 gene product.[77] Olaparib is combined with chemotherapeutics to inhibit single-strand break repair induced by DNA damage caused by the co-administered chemotherapy. Tumor cells relying on this residual DNA repair mechanism are unable to repair the damage and hence are not able to survive and proliferate, whereas normal cells can repair the damage with the functioning homologous recombination mechanism.
Many other drugs for use against other residual DNA repair mechanisms commonly found in cancer are currently under investigation. However, synthetic lethality therapeutic approaches have been questioned due to emerging evidence of acquired resistance, achieved through rewiring of DNA damage response pathways and reversion of previously inhibited defects.[78]
DNA repair defects in cancer[edit]
It has become apparent over the past several years that the DNA damage response acts as a barrier to the malignant transformation of preneoplastic cells.[79] Previous studies have shown an elevated DNA damage response in cell-culture models with oncogene activation[80] and preneoplastic colon adenomas.[81] DNA damage response mechanisms trigger cell-cycle arrest, and attempt to repair DNA lesions or promote cell death/senescence if repair is not possible. Replication stress is observed in preneoplastic cells due to increased proliferation signals from oncogenic mutations. Replication stress is characterized by: increased replication initiation/origin firing; increased transcription and collisions of transcription-replication complexes; nucleotide deficiency; increase in reactive oxygen species (ROS).[82]
Replication stress, along with the selection for inactivating mutations in DNA damage response genes in the evolution of the tumor,[83] leads to downregulation and/or loss of some DNA damage response mechanisms, and hence loss of DNA repair and/or senescence/programmed cell death. In experimental mouse models, loss of DNA damage response-mediated cell senescence was observed after using a short hairpin RNA (shRNA) to inhibit the double-strand break response kinase ataxia telangiectasia (ATM), leading to increased tumor size and invasiveness.[81] Humans born with inherited defects in DNA repair mechanisms (for example, Li-Fraumeni syndrome) have a higher cancer risk.[84]
The prevalence of DNA damage response mutations differs across cancer types; for example, 30% of breast invasive carcinomas have mutations in genes involved in homologous recombination.[79] In cancer, downregulation is observed across all DNA damage response mechanisms (base excision repair (BER), nucleotide excision repair (NER), DNA mismatch repair (MMR), homologous recombination repair (HR), non-homologous end joining (NHEJ) and translesion DNA synthesis (TLS).[85] As well as mutations to DNA damage repair genes, mutations also arise in the genes responsible for arresting the cell cycle to allow sufficient time for DNA repair to occur, and some genes are involved in both DNA damage repair and cell cycle checkpoint control, for example ATM and checkpoint kinase 2 (CHEK2) – a tumor suppressor that is often absent or downregulated in non-small cell lung cancer.[86]
Genes involved in DNA damage response pathways and frequently mutated in cancer (HR = homologous recombination; NHEJ = non-homologous end joining; SSA = single-strand annealing; FA = fanconi anemia pathway; BER = base excision repair; NER = nucleotide excision repair; MMR = mismatch repair)
| HR | NHEJ | SSA | FA | BER | NER | MMR | |
|---|---|---|---|---|---|---|---|
| ATM | |||||||
| ATR | |||||||
| PAXIP | |||||||
| RPA | |||||||
| BRCA1 | |||||||
| BRCA2 | |||||||
| RAD51 | |||||||
| RFC | |||||||
| XRCC1 | |||||||
| PCNA | |||||||
| PARP1 | |||||||
| ERCC1 | |||||||
| MSH3 |
Epigenetic DNA repair defects in cancer[edit]
Classically, cancer has been viewed as a set of diseases that are driven by progressive genetic abnormalities that include mutations in tumour-suppressor genes and oncogenes, and chromosomal aberrations. However, it has become apparent that cancer is also driven by
epigenetic alterations.[87]
Epigenetic alterations refer to functionally relevant modifications to the genome that do not involve a change in the nucleotide sequence. Examples of such modifications are changes in DNA methylation (hypermethylation and hypomethylation) and histone modification,[88] changes in chromosomal architecture (caused by inappropriate expression of proteins such as HMGA2 or HMGA1)[89] and changes caused by microRNAs. Each of these epigenetic alterations serves to regulate gene expression without altering the underlying DNA sequence. These changes usually remain through cell divisions, last for multiple cell generations, and can be considered to be epimutations (equivalent to mutations).
While large numbers of epigenetic alterations are found in cancers, the epigenetic alterations in DNA repair genes, causing reduced expression of DNA repair proteins, appear to be particularly important. Such alterations are thought to occur early in progression to cancer and to be a likely cause of the genetic instability characteristic of cancers.[90][91][92]
Reduced expression of DNA repair genes causes deficient DNA repair. When DNA repair is deficient DNA damages remain in cells at a higher than usual level and these excess damages cause increased frequencies of mutation or epimutation. Mutation rates increase substantially in cells defective in DNA mismatch repair[93][94] or in homologous recombinational repair (HRR).[95] Chromosomal rearrangements and aneuploidy also increase in HRR defective cells.[96]
Higher levels of DNA damage not only cause increased mutation, but also cause increased epimutation. During repair of DNA double strand breaks, or repair of other DNA damages, incompletely cleared sites of repair can cause epigenetic gene silencing.[97][98]
Deficient expression of DNA repair proteins due to an inherited mutation can cause increased risk of cancer. Individuals with an inherited impairment in any of 34 DNA repair genes (see article DNA repair-deficiency disorder) have an increased risk of cancer, with some defects causing up to a 100% lifetime chance of cancer (e.g. p53 mutations).[99] However, such germline mutations (which cause highly penetrant cancer syndromes) are the cause of only about 1 percent of cancers.[100]
Frequencies of epimutations in DNA repair genes[edit]
A chart of common DNA damaging agents, examples of lesions they cause in DNA, and pathways used to repair these lesions. Also shown are many of the genes in these pathways, an indication of which genes are epigenetically regulated to have reduced (or increased) expression in various cancers. It also shows genes in the error-prone microhomology-mediated end joining pathway with increased expression in various cancers.
Deficiencies in DNA repair enzymes are occasionally caused by a newly arising somatic mutation in a DNA repair gene, but are much more frequently caused by epigenetic alterations that reduce or silence expression of DNA repair genes. For example, when 113 colorectal cancers were examined in sequence, only four had a missense mutation in the DNA repair gene MGMT, while the majority had reduced MGMT expression due to methylation of the MGMT promoter region (an epigenetic alteration).[101] Five different studies found that between 40% and 90% of colorectal cancers have reduced MGMT expression due to methylation of the MGMT promoter region.[102][103][104][105][106]
Similarly, out of 119 cases of mismatch repair-deficient colorectal cancers that lacked DNA repair gene PMS2 expression, PMS2 was deficient in 6 due to mutations in the PMS2 gene, while in 103 cases PMS2 expression was deficient because its pairing partner MLH1 was repressed due to promoter methylation (PMS2 protein is unstable in the absence of MLH1).[107] In the other 10 cases, loss of PMS2 expression was likely due to epigenetic overexpression of the microRNA, miR-155, which down-regulates MLH1.[108]
In a further example, epigenetic defects were found in various cancers (e.g. breast, ovarian, colorectal and head and neck). Two or three deficiencies in the expression of ERCC1, XPF or PMS2 occur simultaneously in the majority of 49 colon cancers evaluated by Facista et al.[109]
The chart in this section shows some frequent DNA damaging agents, examples of DNA lesions they cause, and the pathways that deal with these DNA damages. At least 169 enzymes are either directly employed in DNA repair or influence DNA repair processes.[110] Of these, 83 are directly employed in repairing the 5 types of DNA damages illustrated in the chart.
Some of the more well studied genes central to these repair processes are shown in the chart. The gene designations shown in red, gray or cyan indicate genes frequently epigenetically altered in various types of cancers. Wikipedia articles on each of the genes highlighted by red, gray or cyan describe the epigenetic alteration(s) and the cancer(s) in which these epimutations are found. Review articles,[111] and broad experimental survey articles[112][113] also document most of these epigenetic DNA repair deficiencies in cancers.
Red-highlighted genes are frequently reduced or silenced by epigenetic mechanisms in various cancers. When these genes have low or absent expression, DNA damages can accumulate. Replication errors past these damages (see translesion synthesis) can lead to increased mutations and, ultimately, cancer. Epigenetic repression of DNA repair genes in accurate DNA repair pathways appear to be central to carcinogenesis.
The two gray-highlighted genes RAD51 and BRCA2, are required for homologous recombinational repair. They are sometimes epigenetically over-expressed and sometimes under-expressed in certain cancers. As indicated in the Wikipedia articles on RAD51 and BRCA2, such cancers ordinarily have epigenetic deficiencies in other DNA repair genes. These repair deficiencies would likely cause increased unrepaired DNA damages. The over-expression of RAD51 and BRCA2 seen in these cancers may reflect selective pressures for compensatory RAD51 or BRCA2 over-expression and increased homologous recombinational repair to at least partially deal with such excess DNA damages. In those cases where RAD51 or BRCA2 are under-expressed, this would itself lead to increased unrepaired DNA damages. Replication errors past these damages (see translesion synthesis) could cause increased mutations and cancer, so that under-expression of RAD51 or BRCA2 would be carcinogenic in itself.
Cyan-highlighted genes are in the microhomology-mediated end joining (MMEJ) pathway and are up-regulated in cancer. MMEJ is an additional error-prone inaccurate repair pathway for double-strand breaks. In MMEJ repair of a double-strand break, an homology of 5–25 complementary base pairs between both paired strands is sufficient to align the strands, but mismatched ends (flaps) are usually present. MMEJ removes the extra nucleotides (flaps) where strands are joined, and then ligates the strands to create an intact DNA double helix. MMEJ almost always involves at least a small deletion, so that it is a mutagenic pathway.[24] FEN1, the flap endonuclease in MMEJ, is epigenetically increased by promoter hypomethylation and is over-expressed in the majority of cancers of the breast,[114] prostate,[115] stomach,[116][117] neuroblastomas,[118] pancreas,[119] and lung.[120] PARP1 is also over-expressed when its promoter region ETS site is epigenetically hypomethylated, and this contributes to progression to endometrial cancer[121] and BRCA-mutated serous ovarian cancer.[122] Other genes in the MMEJ pathway are also over-expressed in a number of cancers (see MMEJ for summary), and are also shown in cyan.
Genome-wide distribution of DNA repair in human somatic cells[edit]
Differential activity of DNA repair pathways across various regions of the human genome causes mutations to be very unevenly distributed within tumor genomes.[123][124] In particular, the gene-rich, early-replicating regions of the human genome exhibit lower mutation frequencies than the gene-poor, late-replicating heterochromatin. One mechanism underlying this involves the histone modification H3K36me3, which can recruit mismatch repair proteins,[125] thereby lowering mutation rates in H3K36me3-marked regions.[126] Another important mechanism concerns nucleotide excision repair, which can be recruited by the transcription machinery, lowering somatic mutation rates in active genes[124] and other open chromatin regions.[127]
Epigenetic alterations due to DNA repair[edit]
Damage to DNA is very common and is constantly being repaired. Epigenetic alterations can accompany DNA repair of oxidative damage or double-strand breaks. In human cells, oxidative DNA damage occurs about 10,000 times a day and DNA double-strand breaks occur about 10 to 50 times a cell cycle in somatic replicating cells (see DNA damage (naturally occurring)). The selective advantage of DNA repair is to allow the cell to survive in the face of DNA damage. The selective advantage of epigenetic alterations that occur with DNA repair is not clear.
Repair of oxidative DNA damage can alter epigenetic markers[edit]
In the steady state (with endogenous damages occurring and being repaired), there are about 2,400 oxidatively damaged guanines that form 8-oxo-2′-deoxyguanosine (8-OHdG) in the average mamalian cell DNA.[128] 8-OHdG constitutes about 5% of the oxidative damages commonly present in DNA.[129] The oxidized guanines do not occur randomly among all guanines in DNA. There is a sequence preference for the guanine at a methylated CpG site (a cytosine followed by guanine along its 5′ → 3′ direction and where the cytosine is methylated (5-mCpG)).[130] A 5-mCpG site has the lowest ionization potential for guanine oxidation.
Initiation of DNA demethylation at a CpG site. In adult somatic cells DNA methylation typically occurs in the context of CpG dinucleotides (CpG sites), forming 5-methylcytosine-pG, or 5mCpG. Reactive oxygen species (ROS) may attack guanine at the dinucleotide site, forming 8-hydroxy-2′-deoxyguanosine (8-OHdG), and resulting in a 5mCp-8-OHdG dinucleotide site. The base excision repair enzyme OGG1 targets 8-OHdG and binds to the lesion without immediate excision. OGG1, present at a 5mCp-8-OHdG site recruits TET1 and TET1 oxidizes the 5mC adjacent to the 8-OHdG. This initiates demethylation of 5mC.[131]
Oxidized guanine has mispairing potential and is mutagenic.[132] Oxoguanine glycosylase (OGG1) is the primary enzyme responsible for the excision of the oxidized guanine during DNA repair. OGG1 finds and binds to an 8-OHdG within a few seconds.[133] However, OGG1 does not immediately excise 8-OHdG. In HeLa cells half maximum removal of 8-OHdG occurs in 30 minutes,[134] and in irradiated mice, the 8-OHdGs induced in the mouse liver are removed with a half-life of 11 minutes.[129]
When OGG1 is present at an oxidized guanine within a methylated CpG site it recruits TET1 to the 8-OHdG lesion (see Figure). This allows TET1 to demethylate an adjacent methylated cytosine. Demethylation of cytosine is an epigenetic alteration.
As an example, when human mammary epithelial cells were treated with H2O2 for six hours, 8-OHdG increased about 3.5-fold in DNA and this caused about 80% demethylation of the 5-methylcytosines in the genome.[131] Demethylation of CpGs in a gene promoter by TET enzyme activity increases transcription of the gene into messenger RNA.[135] In cells treated with H2O2, one particular gene was examined, BACE1.[131] The methylation level of the BACE1 CpG island was reduced (an epigenetic alteration) and this allowed about 6.5 fold increase of expression of BACE1 messenger RNA.
While six-hour incubation with H2O2 causes considerable demethylation of 5-mCpG sites, shorter times of H2O2 incubation appear to promote other epigenetic alterations. Treatment of cells with H2O2 for 30 minutes causes the mismatch repair protein heterodimer MSH2-MSH6 to recruit DNA methyltransferase 1 (DNMT1) to sites of some kinds of oxidative DNA damage.[136] This could cause increased methylation of cytosines (epigenetic alterations) at these locations.
Jiang et al.[137] treated HEK 293 cells with agents causing oxidative DNA damage, (potassium bromate (KBrO3) or potassium chromate (K2CrO4)). Base excision repair (BER) of oxidative damage occurred with the DNA repair enzyme polymerase beta localizing to oxidized guanines. Polymerase beta is the main human polymerase in short-patch BER of oxidative DNA damage. Jiang et al.[137] also found that polymerase beta recruited the DNA methyltransferase protein DNMT3b to BER repair sites. They then evaluated the methylation pattern at the single nucleotide level in a small region of DNA including the promoter region and the early transcription region of the BRCA1 gene. Oxidative DNA damage from bromate modulated the DNA methylation pattern (caused epigenetic alterations) at CpG sites within the region of DNA studied. In untreated cells, CpGs located at −189, −134, −29, −19, +16, and +19 of the BRCA1 gene had methylated cytosines (where numbering is from the messenger RNA transcription start site, and negative numbers indicate nucleotides in the upstream promoter region). Bromate treatment-induced oxidation resulted in the loss of cytosine methylation at −189, −134, +16 and +19 while also leading to the formation of new methylation at the CpGs located at −80, −55, −21 and +8 after DNA repair was allowed.
Homologous recombinational repair alters epigenetic markers[edit]
At least four articles report the recruitment of DNA methyltransferase 1 (DNMT1) to sites of DNA double-strand breaks.[138][139][140][141] During homologous recombinational repair (HR) of the double-strand break, the involvement of DNMT1 causes the two repaired strands of DNA to have different levels of methylated cytosines. One strand becomes frequently methylated at about 21 CpG sites downstream of the repaired double-strand break. The other DNA strand loses methylation at about six CpG sites that were previously methylated downstream of the double-strand break, as well as losing methylation at about five CpG sites that were previously methylated upstream of the double-strand break. When the chromosome is replicated, this gives rise to one daughter chromosome that is heavily methylated downstream of the previous break site and one that is unmethylated in the region both upstream and downstream of the previous break site. With respect to the gene that was broken by the double-strand break, half of the progeny cells express that gene at a high level and in the other half of the progeny cells expression of that gene is repressed. When clones of these cells were maintained for three years, the new methylation patterns were maintained over that time period.[142]
In mice with a CRISPR-mediated homology-directed recombination insertion in their genome there were a large number of increased methylations of CpG sites within the double-strand break-associated insertion.[143]
Non-homologous end joining can cause some epigenetic marker alterations[edit]
Non-homologous end joining (NHEJ) repair of a double-strand break can cause a small number of demethylations of pre-existing cytosine DNA methylations downstream of the repaired double-strand break.[139] Further work by Allen et al.[144] showed that NHEJ of a DNA double-strand break in a cell could give rise to some progeny cells having repressed expression of the gene harboring the initial double-strand break and some progeny having high expression of that gene due to epigenetic alterations associated with NHEJ repair. The frequency of epigenetic alterations causing repression of a gene after an NHEJ repair of a DNA double-strand break in that gene may be about 0.9%.[140]
Evolution[edit]
The basic processes of DNA repair are highly conserved among both prokaryotes and eukaryotes and even among bacteriophages (viruses which infect bacteria); however, more complex organisms with more complex genomes have correspondingly more complex repair mechanisms.[145] The ability of a large number of protein structural motifs to catalyze relevant chemical reactions has played a significant role in the elaboration of repair mechanisms during evolution. For an extremely detailed review of hypotheses relating to the evolution of DNA repair, see.[146]
The fossil record indicates that single-cell life began to proliferate on the planet at some point during the Precambrian period, although exactly when recognizably modern life first emerged is unclear. Nucleic acids became the sole and universal means of encoding genetic information, requiring DNA repair mechanisms that in their basic form have been inherited by all extant life forms from their common ancestor. The emergence of Earth’s oxygen-rich atmosphere (known as the «oxygen catastrophe») due to photosynthetic organisms, as well as the presence of potentially damaging free radicals in the cell due to oxidative phosphorylation, necessitated the evolution of DNA repair mechanisms that act specifically to counter the types of damage induced by oxidative stress.
Rate of evolutionary change[edit]
On some occasions, DNA damage is not repaired or is repaired by an error-prone mechanism that results in a change from the original sequence. When this occurs, mutations may propagate into the genomes of the cell’s progeny. Should such an event occur in a germ line cell that will eventually produce a gamete, the mutation has the potential to be passed on to the organism’s offspring. The rate of evolution in a particular species (or, in a particular gene) is a function of the rate of mutation. As a consequence, the rate and accuracy of DNA repair mechanisms have an influence over the process of evolutionary change.[147] DNA damage protection and repair does not influence the rate of adaptation by gene regulation and by recombination and selection of alleles. On the other hand, DNA damage repair and protection does influence the rate of accumulation of irreparable, advantageous, code expanding, inheritable mutations, and slows down the evolutionary mechanism for expansion of the genome of organisms with new functionalities. The tension between evolvability and mutation repair and protection needs further investigation.
Technology[edit]
A technology named clustered regularly interspaced short palindromic repeat (shortened to CRISPR-Cas9) was discovered in 2012. The new technology allows anyone with molecular biology training to alter the genes of any species with precision, by inducing DNA damage at a specific point and then altering DNA repair mechanisms to insert new genes.[148] It is cheaper, more efficient, and more precise than other technologies. With the help of CRISPR–Cas9, parts of a genome can be edited by scientists by removing, adding, or altering parts in a DNA sequence.
See also[edit]
- Accelerated aging disease
- Aging DNA
- Cell cycle
- DNA damage (naturally occurring)
- DNA damage theory of aging
- DNA replication
- Direct DNA damage
- Gene therapy
- Human mitochondrial genetics
- Indirect DNA damage
- Life extension
- Progeria
- REPAIRtoire
- Senescence
- SiDNA
- The scientific journal DNA Repair under Mutation Research
References[edit]
- ^ «Nature Reviews Series: DNA damage». Nature Reviews Molecular Cell Biology. 5 July 2017. Retrieved 7 November 2018.
- ^ a b Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J (2004). Molecular Biology of the Cell (5th ed.). New York: WH Freeman. p. 963.
- ^ a b Acharya PV (1971). «The isolation and partial characterization of age-correlated oligo-deoxyribo-ribonucleotides with covalently linked aspartyl-glutamyl polypeptides». Johns Hopkins Medical Journal. Supplement (1): 254–60. PMID 5055816.
- ^ a b Bjorksten J, Acharya PV, Ashman S, Wetlaufer DB (July 1971). «Gerogenic fractions in the tritiated rat». Journal of the American Geriatrics Society. 19 (7): 561–74. doi:10.1111/j.1532-5415.1971.tb02577.x. PMID 5106728. S2CID 33154242.
- ^ Browner WS, Kahn AJ, Ziv E, Reiner AP, Oshima J, Cawthon RM, et al. (December 2004). «The genetics of human longevity». The American Journal of Medicine. 117 (11): 851–60. CiteSeerX 10.1.1.556.6874. doi:10.1016/j.amjmed.2004.06.033. PMID 15589490.
- ^ Broad WJ (7 October 2015). «Nobel Prize in Chemistry Awarded to Tomas Lindahl, Paul Modrich and Aziz Sancar for DNA Studies». The New York Times. Retrieved 7 October 2015.
- ^ Staff (7 October 2015). «The Nobel Prize in Chemistry 2015 – DNA repair – providing chemical stability for life» (PDF). Nobel Prize. Retrieved 7 October 2015.
- ^ Roulston A, Marcellus RC, Branton PE (1999). «Viruses and apoptosis». Annual Review of Microbiology. 53: 577–628. doi:10.1146/annurev.micro.53.1.577. PMID 10547702.
- ^ Madigan MT, Martino JM (2006). Brock Biology of Microorganisms (11th ed.). Pearson. p. 136. ISBN 978-0-13-196893-6.
- ^ Ohta T, Tokishita SI, Mochizuki K, Kawase J, Sakahira M, Yamagata H (2006). «UV Sensitivity and Mutagenesis of the Extremely Thermophilic Eubacterium Thermus thermophilus HB27». Genes and Environment. 28 (2): 56–61. doi:10.3123/jemsge.28.56.
- ^ Tanaka T, Halicka HD, Huang X, Traganos F, Darzynkiewicz Z (September 2006). «Constitutive histone H2AX phosphorylation and ATM activation, the reporters of DNA damage by endogenous oxidants». Cell Cycle. 5 (17): 1940–45. doi:10.4161/cc.5.17.3191. PMC 3488278. PMID 16940754.
- ^ Braig M, Schmitt CA (March 2006). «Oncogene-induced senescence: putting the brakes on tumor development». Cancer Research. 66 (6): 2881–84. doi:10.1158/0008-5472.CAN-05-4006. PMID 16540631.
- ^ Lynch MD (February 2006). «How does cellular senescence prevent cancer?». DNA and Cell Biology. 25 (2): 69–78. doi:10.1089/dna.2006.25.69. PMID 16460230.
- ^ Campisi J, d’Adda di Fagagna F (September 2007). «Cellular senescence: when bad things happen to good cells». Nature Reviews. Molecular Cell Biology. 8 (9): 729–40. doi:10.1038/nrm2233. PMID 17667954. S2CID 15664931.
- ^ a b c Best BP (June 2009). «Nuclear DNA damage as a direct cause of aging» (PDF). Rejuvenation Research. 12 (3): 199–208. CiteSeerX 10.1.1.318.738. doi:10.1089/rej.2009.0847. PMID 19594328. Archived from the original (PDF) on 15 November 2017. Retrieved 29 September 2009.
- ^ Sancar A (June 2003). «Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors». Chemical Reviews. 103 (6): 2203–37. doi:10.1021/cr0204348. PMID 12797829.
- ^ Lucas-Lledó JI, Lynch M (May 2009). «Evolution of mutation rates: phylogenomic analysis of the photolyase/cryptochrome family». Molecular Biology and Evolution. 26 (5): 1143–53. doi:10.1093/molbev/msp029. PMC 2668831. PMID 19228922.
- ^ a b c Watson JD, Baker TA, Bell SP, Gann A, Levine M, Losick R (2004). Molecular Biology of the Gene (5th ed.). Pearson Benjamin Cummings; CSHL Press. Ch. 9, 10. OCLC 936762772.
- ^ Volkert MR (1988). «Adaptive response of Escherichia coli to alkylation damage». Environmental and Molecular Mutagenesis. 11 (2): 241–55. doi:10.1002/em.2850110210. PMID 3278898. S2CID 24722637.
- ^ a b c Willey J, Sherwood L, Woolverton C (2014). Prescott’s Microbiology. New York: McGraw Hill. p. 381. ISBN 978-0-07-3402-40-6.
- ^ Russell P (2018). i Genetics. Chennai: Pearson. p. 186. ISBN 978-93-325-7162-4.
- ^ a b c d Reardon JT, Sancar A (2006). «Purification and characterization of Escherichia coli and human nucleotide excision repair enzyme systems». Methods in Enzymology. 408: 189–213. doi:10.1016/S0076-6879(06)08012-8. ISBN 9780121828134. PMID 16793370.
- ^ Berg M, Tymoczko J, Stryer L (2012). Biochemistry 7th edition. New York: W.H. Freeman and Company. p. 840. ISBN 9781429229364.
- ^ a b Liang L, Deng L, Chen Y, Li GC, Shao C, Tischfield JA (September 2005). «Modulation of DNA end joining by nuclear proteins». The Journal of Biological Chemistry. 280 (36): 31442–49. doi:10.1074/jbc.M503776200. PMID 16012167.
- ^ Wilson TE, Grawunder U, Lieber MR (July 1997). «Yeast DNA ligase IV mediates non-homologous DNA end joining». Nature. 388 (6641): 495–98. Bibcode:1997Natur.388..495W. doi:10.1038/41365. PMID 9242411. S2CID 4422938.
- ^ Moore JK, Haber JE (May 1996). «Cell cycle and genetic requirements of two pathways of nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae». Molecular and Cellular Biology. 16 (5): 2164–73. doi:10.1128/mcb.16.5.2164. PMC 231204. PMID 8628283.
- ^ Boulton SJ, Jackson SP (September 1996). «Saccharomyces cerevisiae Ku70 potentiates illegitimate DNA double-strand break repair and serves as a barrier to error-prone DNA repair pathways». The EMBO Journal. 15 (18): 5093–103. doi:10.1002/j.1460-2075.1996.tb00890.x. PMC 452249. PMID 8890183.
- ^ Wilson TE, Lieber MR (August 1999). «Efficient processing of DNA ends during yeast nonhomologous end joining. Evidence for a DNA polymerase beta (Pol4)-dependent pathway». The Journal of Biological Chemistry. 274 (33): 23599–609. doi:10.1074/jbc.274.33.23599. PMID 10438542.
- ^ Budman J, Chu G (February 2005). «Processing of DNA for nonhomologous end-joining by cell-free extract». The EMBO Journal. 24 (4): 849–60. doi:10.1038/sj.emboj.7600563. PMC 549622. PMID 15692565.
- ^ Wang H, Perrault AR, Takeda Y, Qin W, Wang H, Iliakis G (September 2003). «Biochemical evidence for Ku-independent backup pathways of NHEJ». Nucleic Acids Research. 31 (18): 5377–88. doi:10.1093/nar/gkg728. PMC 203313. PMID 12954774.
- ^ Jung D, Alt FW (January 2004). «Unraveling V(D)J recombination; insights into gene regulation». Cell. 116 (2): 299–311. doi:10.1016/S0092-8674(04)00039-X. PMID 14744439. S2CID 16890458.
- ^ a b Truong LN, Li Y, Shi LZ, Hwang PY, He J, Wang H, et al. (May 2013). «Microhomology-mediated End Joining and Homologous Recombination share the initial end resection step to repair DNA double-strand breaks in mammalian cells». Proceedings of the National Academy of Sciences of the United States of America. 110 (19): 7720–25. Bibcode:2013PNAS..110.7720T. doi:10.1073/pnas.1213431110. PMC 3651503. PMID 23610439.
- ^ Sharma S, Javadekar SM, Pandey M, Srivastava M, Kumari R, Raghavan SC (March 2015). «Homology and enzymatic requirements of microhomology-dependent alternative end joining». Cell Death & Disease. 6 (3): e1697. doi:10.1038/cddis.2015.58. PMC 4385936. PMID 25789972.
- ^ Decottignies A (2013). «Alternative end-joining mechanisms: a historical perspective». Frontiers in Genetics. 4: 48. doi:10.3389/fgene.2013.00048. PMC 3613618. PMID 23565119.
- ^ Zahradka K, Slade D, Bailone A, Sommer S, Averbeck D, Petranovic M, et al. (October 2006). «Reassembly of shattered chromosomes in Deinococcus radiodurans». Nature. 443 (7111): 569–73. Bibcode:2006Natur.443..569Z. doi:10.1038/nature05160. PMID 17006450. S2CID 4412830.
- ^ Stenerlöw B, Karlsson KH, Cooper B, Rydberg B. «Measurement of prompt DNA double-strand breaks in mammalian cells without including heat-labile sites: results for cells deficient in nonhomologous end joining». Radiat Res. 2003 Apr;159(4):502–10. doi:10.1667/0033-7587(2003)159[0502:mopdds2.0.co;2] PMID 12643795.
- ^ Waters LS, Minesinger BK, Wiltrout ME, D’Souza S, Woodruff RV, Walker GC (March 2009). «Eukaryotic translesion polymerases and their roles and regulation in DNA damage tolerance». Microbiology and Molecular Biology Reviews. 73 (1): 134–54. doi:10.1128/MMBR.00034-08. PMC 2650891. PMID 19258535.
- ^ Colis LC, Raychaudhury P, Basu AK (August 2008). «Mutational specificity of gamma-radiation-induced guanine-thymine and thymine-guanine intrastrand cross-links in mammalian cells and translesion synthesis past the guanine-thymine lesion by human DNA polymerase eta». Biochemistry. 47 (31): 8070–79. doi:10.1021/bi800529f. PMC 2646719. PMID 18616294.
- ^ Raychaudhury P, Basu AK (March 2011). «Genetic requirement for mutagenesis of the G[8,5-Me]T cross-link in Escherichia coli: DNA polymerases IV and V compete for error-prone bypass». Biochemistry. 50 (12): 2330–38. doi:10.1021/bi102064z. PMC 3062377. PMID 21302943.
- ^ «Translesion Synthesis». Research.chem.psu.edu. Archived from the original on 10 March 2012. Retrieved 14 August 2012.
- ^ Wang Z (July 2001). «Translesion synthesis by the UmuC family of DNA polymerases». Mutation Research. 486 (2): 59–70. doi:10.1016/S0921-8777(01)00089-1. PMID 11425512.
- ^ a b c Friedberg EC, Walker GC, Siede W, Wood RD, Schultz RA, Ellenberger T. (2006). DNA Repair and Mutagenesis, part 3. ASM Press. 2nd ed.
- ^ Liu B, Yip RK, Zhou Z (November 2012). «Chromatin remodeling, DNA damage repair and aging». Current Genomics. 13 (7): 533–47. doi:10.2174/138920212803251373. PMC 3468886. PMID 23633913.
- ^ Halicka HD, Zhao H, Podhorecka M, Traganos F, Darzynkiewicz Z (July 2009). «Cytometric detection of chromatin relaxation, an early reporter of DNA damage response». Cell Cycle. 8 (14): 2233–37. doi:10.4161/cc.8.14.8984. PMC 3856216. PMID 19502789.
- ^ a b c Sellou H, Lebeaupin T, Chapuis C, Smith R, Hegele A, Singh HR, et al. (December 2016). «The poly(ADP-ribose)-dependent chromatin remodeler Alc1 induces local chromatin relaxation upon DNA damage». Molecular Biology of the Cell. 27 (24): 3791–99. doi:10.1091/mbc.E16-05-0269. PMC 5170603. PMID 27733626.
- ^ a b Van Meter M, Simon M, Tombline G, May A, Morello TD, Hubbard BP, et al. (September 2016). «JNK Phosphorylates SIRT6 to Stimulate DNA Double-Strand Break Repair in Response to Oxidative Stress by Recruiting PARP1 to DNA Breaks». Cell Reports. 16 (10): 2641–50. doi:10.1016/j.celrep.2016.08.006. PMC 5089070. PMID 27568560.
- ^ a b Haince JF, McDonald D, Rodrigue A, Déry U, Masson JY, Hendzel MJ, Poirier GG (January 2008). «PARP1-dependent kinetics of recruitment of MRE11 and NBS1 proteins to multiple DNA damage sites». The Journal of Biological Chemistry. 283 (2): 1197–208. doi:10.1074/jbc.M706734200. PMID 18025084.
- ^ a b c Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM (March 1998). «DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139». The Journal of Biological Chemistry. 273 (10): 5858–68. doi:10.1074/jbc.273.10.5858. PMID 9488723.
- ^ Mailand N, Bekker-Jensen S, Faustrup H, Melander F, Bartek J, Lukas C, Lukas J (November 2007). «RNF8 ubiquitylates histones at DNA double-strand breaks and promotes assembly of repair proteins». Cell. 131 (5): 887–900. doi:10.1016/j.cell.2007.09.040. PMID 18001824. S2CID 14232192.
- ^ Luijsterburg MS, Acs K, Ackermann L, Wiegant WW, Bekker-Jensen S, Larsen DH, et al. (May 2012). «A new non-catalytic role for ubiquitin ligase RNF8 in unfolding higher-order chromatin structure». The EMBO Journal. 31 (11): 2511–27. doi:10.1038/emboj.2012.104. PMC 3365417. PMID 22531782.
- ^ a b Luijsterburg MS, Goedhart J, Moser J, Kool H, Geverts B, Houtsmuller AB, et al. (August 2007). «Dynamic in vivo interaction of DDB2 E3 ubiquitin ligase with UV-damaged DNA is independent of damage-recognition protein XPC». Journal of Cell Science. 120 (Pt 15): 2706–16. doi:10.1242/jcs.008367. PMID 17635991.
- ^ a b Pines A, Vrouwe MG, Marteijn JA, Typas D, Luijsterburg MS, Cansoy M, et al. (October 2012). «PARP1 promotes nucleotide excision repair through DDB2 stabilization and recruitment of ALC1». The Journal of Cell Biology. 199 (2): 235–49. doi:10.1083/jcb.201112132. PMC 3471223. PMID 23045548.
- ^ Jazayeri A, Falck J, Lukas C, Bartek J, Smith GC, Lukas J, Jackson SP (January 2006). «ATM- and cell cycle-dependent regulation of ATR in response to DNA double-strand breaks». Nature Cell Biology. 8 (1): 37–45. doi:10.1038/ncb1337. PMID 16327781. S2CID 9797133.
- ^ Bakkenist CJ, Kastan MB (January 2003). «DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation». Nature. 421 (6922): 499–506. Bibcode:2003Natur.421..499B. doi:10.1038/nature01368. PMID 12556884. S2CID 4403303.
- ^ Wei Q, Li L, Chen D (2007). DNA Repair, Genetic Instability, and Cancer. World Scientific. ISBN 978-981-270-014-8.[page needed]
- ^ Schonthal AH (2004). Checkpoint Controls and Cancer. Humana Press. ISBN 978-1-58829-500-2.[page needed]
- ^ Gartel AL, Tyner AL (June 2002). «The role of the cyclin-dependent kinase inhibitor p21 in apoptosis». Molecular Cancer Therapeutics. 1 (8): 639–49. PMID 12479224.
- ^ Janion C (2001). «Some aspects of the SOS response system—a critical survey». Acta Biochimica Polonica. 48 (3): 599–610. doi:10.18388/abp.2001_3894. PMID 11833768.
- ^ a b Erill I, Campoy S, Barbé J (November 2007). «Aeons of distress: an evolutionary perspective on the bacterial SOS response». FEMS Microbiology Reviews. 31 (6): 637–56. doi:10.1111/j.1574-6976.2007.00082.x. PMID 17883408.
- ^ Schlacher K, Pham P, Cox MM, Goodman MF (February 2006). «Roles of DNA polymerase V and RecA protein in SOS damage-induced mutation». Chemical Reviews. 106 (2): 406–19. doi:10.1021/cr0404951. PMID 16464012.
- ^ Fry RC, Begley TJ, Samson LD (2004). «Genome-wide responses to DNA-damaging agents». Annual Review of Microbiology. 59: 357–77. doi:10.1146/annurev.micro.59.031805.133658. PMID 16153173.
- ^ Espejel S, Martín M, Klatt P, Martín-Caballero J, Flores JM, Blasco MA (May 2004). «Shorter telomeres, accelerated ageing and increased lymphoma in DNA-PKcs-deficient mice». EMBO Reports. 5 (5): 503–09. doi:10.1038/sj.embor.7400127. PMC 1299048. PMID 15105825.
- ^ de Boer J, Andressoo JO, de Wit J, Huijmans J, Beems RB, van Steeg H, et al. (May 2002). «Premature aging in mice deficient in DNA repair and transcription». Science. 296 (5571): 1276–79. Bibcode:2002Sci…296.1276D. doi:10.1126/science.1070174. PMID 11950998. S2CID 41930529.
- ^ Dollé ME, Busuttil RA, Garcia AM, Wijnhoven S, van Drunen E, Niedernhofer LJ, et al. (April 2006). «Increased genomic instability is not a prerequisite for shortened lifespan in DNA repair deficient mice». Mutation Research. 596 (1–2): 22–35. doi:10.1016/j.mrfmmm.2005.11.008. PMID 16472827.
- ^ Kobayashi Y, Narumi I, Satoh K, Funayama T, Kikuchi M, Kitayama S, Watanabe H (November 2004). «Radiation response mechanisms of the extremely radioresistant bacterium Deinococcus radiodurans». Uchu Seibutsu Kagaku. 18 (3): 134–35. PMID 15858357.
- ^ Spindler SR (September 2005). «Rapid and reversible induction of the longevity, anticancer and genomic effects of caloric restriction». Mechanisms of Ageing and Development. 126 (9): 960–66. doi:10.1016/j.mad.2005.03.016. PMID 15927235. S2CID 7067036.
- ^ Halicka HD, Zhao H, Li J, Lee YS, Hsieh TC, Wu JM, Darzynkiewicz Z (December 2012). «Potential anti-aging agents suppress the level of constitutive mTOR- and DNA damage- signaling». Aging. 4 (12): 952–65. doi:10.18632/aging.100521. PMC 3615161. PMID 23363784.
- ^ Tissenbaum HA, Guarente L (March 2001). «Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans». Nature. 410 (6825): 227–30. Bibcode:2001Natur.410..227T. doi:10.1038/35065638. PMID 11242085. S2CID 4356885.
- ^ Cohen HY, Miller C, Bitterman KJ, Wall NR, Hekking B, Kessler B, et al. (July 2004). «Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase». Science. 305 (5682): 390–92. Bibcode:2004Sci…305..390C. doi:10.1126/science.1099196. PMID 15205477. S2CID 33503081.
- ^ Cabelof DC, Yanamadala S, Raffoul JJ, Guo Z, Soofi A, Heydari AR (March 2003). «Caloric restriction promotes genomic stability by induction of base excision repair and reversal of its age-related decline». DNA Repair. 2 (3): 295–307. doi:10.1016/S1568-7864(02)00219-7. PMID 12547392.
- ^ Stuart JA, Karahalil B, Hogue BA, Souza-Pinto NC, Bohr VA (March 2004). «Mitochondrial and nuclear DNA base excision repair are affected differently by caloric restriction». FASEB Journal. 18 (3): 595–97. doi:10.1096/fj.03-0890fje. PMID 14734635. S2CID 43118901.
- ^ Walker DW, McColl G, Jenkins NL, Harris J, Lithgow GJ (May 2000). «Evolution of lifespan in C. elegans». Nature. 405 (6784): 296–97. doi:10.1038/35012693. PMID 10830948. S2CID 4402039.
- ^ Johnson G (28 December 2010). «Unearthing Prehistoric Tumors, and Debate». The New York Times.
If we lived long enough, sooner or later we all would get cancer.
- ^ Alberts B, Johnson A, Lewis J, et al. (2002). «The Preventable Causes of Cancer». Molecular biology of the cell (4th ed.). New York: Garland Science. ISBN 978-0-8153-4072-0.
A certain irreducible background incidence of cancer is to be expected regardless of circumstances: mutations can never be absolutely avoided, because they are an inescapable consequence of fundamental limitations on the accuracy of DNA replication, as discussed in Chapter 5. If a human could live long enough, it is inevitable that at least one of his or her cells would eventually accumulate a set of mutations sufficient for cancer to develop.
- ^ Friedenson B (August 2007). «The BRCA1/2 pathway prevents hematologic cancers in addition to breast and ovarian cancers». BMC Cancer. 7: 152. doi:10.1186/1471-2407-7-152. PMC 1959234. PMID 17683622.
- ^ Gavande NS, VanderVere-Carozza PS, Hinshaw HD, Jalal SI, Sears CR, Pawelczak KS, Turchi JJ (April 2016). «DNA repair targeted therapy: The past or future of cancer treatment?». Pharmacology & Therapeutics. 160: 65–83. doi:10.1016/j.pharmthera.2016.02.003. PMC 4811676. PMID 26896565.
- ^ Bryant HE, Schultz N, Thomas HD, Parker KM, Flower D, Lopez E, et al. (April 2005). «Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase». Nature. 434 (7035): 913–17. Bibcode:2005Natur.434..913B. doi:10.1038/nature03443. PMID 15829966. S2CID 4391043.
- ^ Goldstein M, Kastan MB (2015). «The DNA damage response: implications for tumor responses to radiation and chemotherapy». Annual Review of Medicine. 66: 129–43. doi:10.1146/annurev-med-081313-121208. PMID 25423595.
- ^ a b Jeggo PA, Pearl LH, Carr AM (January 2016). «DNA repair, genome stability and cancer: a historical perspective» (PDF). Nature Reviews. Cancer. 16 (1): 35–42. doi:10.1038/nrc.2015.4. PMID 26667849. S2CID 14941857.
- ^ Bartkova J, Horejsí Z, Koed K, Krämer A, Tort F, Zieger K, et al. (April 2005). «DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis». Nature. 434 (7035): 864–70. Bibcode:2005Natur.434..864B. doi:10.1038/nature03482. PMID 15829956. S2CID 4398393.
- ^ a b Bartkova J, Rezaei N, Liontos M, Karakaidos P, Kletsas D, Issaeva N, et al. (November 2006). «Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints». Nature. 444 (7119): 633–37. Bibcode:2006Natur.444..633B. doi:10.1038/nature05268. PMID 17136093. S2CID 4406956.
- ^ Gaillard H, García-Muse T, Aguilera A (May 2015). «Replication stress and cancer». Nature Reviews. Cancer. 15 (5): 276–89. doi:10.1038/nrc3916. hdl:10261/123721. PMID 25907220. S2CID 11342123.
- ^ Halazonetis TD, Gorgoulis VG, Bartek J (March 2008). «An oncogene-induced DNA damage model for cancer development». Science. 319 (5868): 1352–55. Bibcode:2008Sci…319.1352H. doi:10.1126/science.1140735. PMID 18323444. S2CID 16426080.
- ^ de Boer J, Hoeijmakers JH (March 2000). «Nucleotide excision repair and human syndromes» (PDF). Carcinogenesis. 21 (3): 453–60. doi:10.1093/carcin/21.3.453. PMID 10688865.
- ^ Broustas CG, Lieberman HB (February 2014). «DNA damage response genes and the development of cancer metastasis». Radiation Research. 181 (2): 111–30. Bibcode:2014RadR..181..111B. doi:10.1667/RR13515.1. PMC 4064942. PMID 24397478.
- ^ Zhang P, Wang J, Gao W, Yuan BZ, Rogers J, Reed E (May 2004). «CHK2 kinase expression is down-regulated due to promoter methylation in non-small cell lung cancer». Molecular Cancer. 3 (4): 14. doi:10.1186/1476-4598-3-14. PMC 419366. PMID 15125777.
- ^ Baylin SB, Ohm JE (February 2006). «Epigenetic gene silencing in cancer – a mechanism for early oncogenic pathway addiction?». Nature Reviews. Cancer. 6 (2): 107–16. doi:10.1038/nrc1799. PMID 16491070. S2CID 2514545.
- ^ Kanwal R, Gupta S (April 2012). «Epigenetic modifications in cancer». Clinical Genetics. 81 (4): 303–11. doi:10.1111/j.1399-0004.2011.01809.x. PMC 3590802. PMID 22082348.
- ^ Baldassarre G, Battista S, Belletti B, Thakur S, Pentimalli F, Trapasso F, et al. (April 2003). «Negative regulation of BRCA1 gene expression by HMGA1 proteins accounts for the reduced BRCA1 protein levels in sporadic breast carcinoma». Molecular and Cellular Biology. 23 (7): 2225–38. doi:10.1128/MCB.23.7.2225-2238.2003. PMC 150734. PMID 12640109.
- ^ Jacinto FV, Esteller M (July 2007). «Mutator pathways unleashed by epigenetic silencing in human cancer». Mutagenesis. 22 (4): 247–53. doi:10.1093/mutage/gem009. PMID 17412712.
- ^ Lahtz C, Pfeifer GP (February 2011). «Epigenetic changes of DNA repair genes in cancer». Journal of Molecular Cell Biology. 3 (1): 51–58. doi:10.1093/jmcb/mjq053. PMC 3030973. PMID 21278452. Epigenetic changes of DNA repair genes in cancer
- ^ Bernstein C, Nfonsam V, Prasad AR, Bernstein H (March 2013). «Epigenetic field defects in progression to cancer». World Journal of Gastrointestinal Oncology. 5 (3): 43–49. doi:10.4251/wjgo.v5.i3.43. PMC 3648662. PMID 23671730.
- ^ Narayanan L, Fritzell JA, Baker SM, Liskay RM, Glazer PM (April 1997). «Elevated levels of mutation in multiple tissues of mice deficient in the DNA mismatch repair gene Pms2». Proceedings of the National Academy of Sciences of the United States of America. 94 (7): 3122–27. Bibcode:1997PNAS…94.3122N. doi:10.1073/pnas.94.7.3122. PMC 20332. PMID 9096356.
- ^ Hegan DC, Narayanan L, Jirik FR, Edelmann W, Liskay RM, Glazer PM (December 2006). «Differing patterns of genetic instability in mice deficient in the mismatch repair genes Pms2, Mlh1, Msh2, Msh3 and Msh6». Carcinogenesis. 27 (12): 2402–08. doi:10.1093/carcin/bgl079. PMC 2612936. PMID 16728433.
- ^ Tutt AN, van Oostrom CT, Ross GM, van Steeg H, Ashworth A (March 2002). «Disruption of Brca2 increases the spontaneous mutation rate in vivo: synergism with ionizing radiation». EMBO Reports. 3 (3): 255–60. doi:10.1093/embo-reports/kvf037. PMC 1084010. PMID 11850397.
- ^ German J (March 1969). «Bloom’s syndrome. I. Genetical and clinical observations in the first twenty-seven patients». American Journal of Human Genetics. 21 (2): 196–227. PMC 1706430. PMID 5770175.
- ^ O’Hagan HM, Mohammad HP, Baylin SB (August 2008). «Double strand breaks can initiate gene silencing and SIRT1-dependent onset of DNA methylation in an exogenous promoter CpG island». PLOS Genetics. 4 (8): e1000155. doi:10.1371/journal.pgen.1000155. PMC 2491723. PMID 18704159.
- ^ Cuozzo C, Porcellini A, Angrisano T, Morano A, Lee B, Di Pardo A, et al. (July 2007). «DNA damage, homology-directed repair, and DNA methylation». PLOS Genetics. 3 (7): e110. doi:10.1371/journal.pgen.0030110. PMC 1913100. PMID 17616978.
- ^ Malkin D (April 2011). «Li-fraumeni syndrome». Genes & Cancer. 2 (4): 475–84. doi:10.1177/1947601911413466. PMC 3135649. PMID 21779515.
- ^ Fearon ER (November 1997). «Human cancer syndromes: clues to the origin and nature of cancer». Science. 278 (5340): 1043–50. Bibcode:1997Sci…278.1043F. doi:10.1126/science.278.5340.1043. PMID 9353177.
- ^ Halford S, Rowan A, Sawyer E, Talbot I, Tomlinson I (June 2005). «O(6)-methylguanine methyltransferase in colorectal cancers: detection of mutations, loss of expression, and weak association with G:C>A:T transitions». Gut. 54 (6): 797–802. doi:10.1136/gut.2004.059535. PMC 1774551. PMID 15888787.
- ^ Shen L, Kondo Y, Rosner GL, Xiao L, Hernandez NS, Vilaythong J, et al. (September 2005). «MGMT promoter methylation and field defect in sporadic colorectal cancer». Journal of the National Cancer Institute. 97 (18): 1330–38. doi:10.1093/jnci/dji275. PMID 16174854.
- ^ Psofaki V, Kalogera C, Tzambouras N, Stephanou D, Tsianos E, Seferiadis K, Kolios G (July 2010). «Promoter methylation status of hMLH1, MGMT, and CDKN2A/p16 in colorectal adenomas». World Journal of Gastroenterology. 16 (28): 3553–60. doi:10.3748/wjg.v16.i28.3553. PMC 2909555. PMID 20653064.
- ^ Lee KH, Lee JS, Nam JH, Choi C, Lee MC, Park CS, et al. (October 2011). «Promoter methylation status of hMLH1, hMSH2, and MGMT genes in colorectal cancer associated with adenoma-carcinoma sequence». Langenbeck’s Archives of Surgery. 396 (7): 1017–26. doi:10.1007/s00423-011-0812-9. PMID 21706233. S2CID 8069716.
- ^ Amatu A, Sartore-Bianchi A, Moutinho C, Belotti A, Bencardino K, Chirico G, et al. (April 2013). «Promoter CpG island hypermethylation of the DNA repair enzyme MGMT predicts clinical response to dacarbazine in a phase II study for metastatic colorectal cancer». Clinical Cancer Research. 19 (8): 2265–72. doi:10.1158/1078-0432.CCR-12-3518. PMID 23422094.
- ^ Mokarram P, Zamani M, Kavousipour S, Naghibalhossaini F, Irajie C, Moradi Sarabi M, Hosseini SV (May 2013). «Different patterns of DNA methylation of the two distinct O6-methylguanine-DNA methyltransferase (O6-MGMT) promoter regions in colorectal cancer». Molecular Biology Reports. 40 (5): 3851–57. doi:10.1007/s11033-012-2465-3. PMID 23271133. S2CID 18733871.
- ^ Truninger K, Menigatti M, Luz J, Russell A, Haider R, Gebbers JO, et al. (May 2005). «Immunohistochemical analysis reveals high frequency of PMS2 defects in colorectal cancer». Gastroenterology. 128 (5): 1160–71. doi:10.1053/j.gastro.2005.01.056. PMID 15887099.
- ^ Valeri N, Gasparini P, Fabbri M, Braconi C, Veronese A, Lovat F, et al. (April 2010). «Modulation of mismatch repair and genomic stability by miR-155». Proceedings of the National Academy of Sciences of the United States of America. 107 (15): 6982–87. Bibcode:2010PNAS..107.6982V. doi:10.1073/pnas.1002472107. PMC 2872463. PMID 20351277.
- ^ Facista A, Nguyen H, Lewis C, Prasad AR, Ramsey L, Zaitlin B, et al. (April 2012). «Deficient expression of DNA repair enzymes in early progression to sporadic colon cancer». Genome Integrity. 3 (1): 3. doi:10.1186/2041-9414-3-3. PMC 3351028. PMID 22494821.
- ^ Human DNA Repair Genes, 15 April 2014, MD Anderson Cancer Center, University of Texas
- ^ Jin B, Robertson KD (2013). «DNA methyltransferases, DNA damage repair, and cancer». Advances in Experimental Medicine and Biology. 754: 3–29. doi:10.1007/978-1-4419-9967-2_1. ISBN 978-1-4419-9966-5. PMC 3707278. PMID 22956494.
- ^ Krishnan K, Steptoe AL, Martin HC, Wani S, Nones K, Waddell N, et al. (February 2013). «MicroRNA-182-5p targets a network of genes involved in DNA repair». RNA. 19 (2): 230–42. doi:10.1261/rna.034926.112. PMC 3543090. PMID 23249749.
- ^ Chaisaingmongkol J, Popanda O, Warta R, Dyckhoff G, Herpel E, Geiselhart L, et al. (December 2012). «Epigenetic screen of human DNA repair genes identifies aberrant promoter methylation of NEIL1 in head and neck squamous cell carcinoma». Oncogene. 31 (49): 5108–16. doi:10.1038/onc.2011.660. PMID 22286769.
- ^ Singh P, Yang M, Dai H, Yu D, Huang Q, Tan W, et al. (November 2008). «Overexpression and hypomethylation of flap endonuclease 1 gene in breast and other cancers». Molecular Cancer Research. 6 (11): 1710–17. doi:10.1158/1541-7786.MCR-08-0269. PMC 2948671. PMID 19010819.
- ^ Lam JS, Seligson DB, Yu H, Li A, Eeva M, Pantuck AJ, et al. (August 2006). «Flap endonuclease 1 is overexpressed in prostate cancer and is associated with a high Gleason score». BJU International. 98 (2): 445–51. doi:10.1111/j.1464-410X.2006.06224.x. PMID 16879693. S2CID 22165252.
- ^ Kim JM, Sohn HY, Yoon SY, Oh JH, Yang JO, Kim JH, et al. (January 2005). «Identification of gastric cancer-related genes using a cDNA microarray containing novel expressed sequence tags expressed in gastric cancer cells». Clinical Cancer Research. 11 (2 Pt 1): 473–82. doi:10.1158/1078-0432.473.11.2. PMID 15701830.
- ^ Wang K, Xie C, Chen D (May 2014). «Flap endonuclease 1 is a promising candidate biomarker in gastric cancer and is involved in cell proliferation and apoptosis». International Journal of Molecular Medicine. 33 (5): 1268–74. doi:10.3892/ijmm.2014.1682. PMID 24590400.
- ^ Krause A, Combaret V, Iacono I, Lacroix B, Compagnon C, Bergeron C, et al. (July 2005). «Genome-wide analysis of gene expression in neuroblastomas detected by mass screening» (PDF). Cancer Letters. 225 (1): 111–20. doi:10.1016/j.canlet.2004.10.035. PMID 15922863. S2CID 44644467.
- ^ Iacobuzio-Donahue CA, Maitra A, Olsen M, Lowe AW, van Heek NT, Rosty C, et al. (April 2003). «Exploration of global gene expression patterns in pancreatic adenocarcinoma using cDNA microarrays». The American Journal of Pathology. 162 (4): 1151–62. doi:10.1016/S0002-9440(10)63911-9. PMC 1851213. PMID 12651607.
- ^ Sato M, Girard L, Sekine I, Sunaga N, Ramirez RD, Kamibayashi C, Minna JD (October 2003). «Increased expression and no mutation of the Flap endonuclease (FEN1) gene in human lung cancer». Oncogene. 22 (46): 7243–46. doi:10.1038/sj.onc.1206977. PMID 14562054.
- ^ Bi FF, Li D, Yang Q (2013). «Hypomethylation of ETS transcription factor binding sites and upregulation of PARP1 expression in endometrial cancer». BioMed Research International. 2013: 946268. doi:10.1155/2013/946268. PMC 3666359. PMID 23762867.
- ^ Bi FF, Li D, Yang Q (February 2013). «Promoter hypomethylation, especially around the E26 transformation-specific motif, and increased expression of poly (ADP-ribose) polymerase 1 in BRCA-mutated serous ovarian cancer». BMC Cancer. 13: 90. doi:10.1186/1471-2407-13-90. PMC 3599366. PMID 23442605.
- ^ Supek F, Lehner B (May 2015). «Differential DNA mismatch repair underlies mutation rate variation across the human genome». Nature. 521 (7550): 81–84. Bibcode:2015Natur.521…81S. doi:10.1038/nature14173. PMC 4425546. PMID 25707793.
- ^ a b Zheng CL, Wang NJ, Chung J, Moslehi H, Sanborn JZ, Hur JS, et al. (November 2014). «Transcription restores DNA repair to heterochromatin, determining regional mutation rates in cancer genomes». Cell Reports. 9 (4): 1228–34. doi:10.1016/j.celrep.2014.10.031. PMC 4254608. PMID 25456125.
- ^ Li F, Mao G, Tong D, Huang J, Gu L, Yang W, Li GM (April 2013). «The histone mark H3K36me3 regulates human DNA mismatch repair through its interaction with MutSα». Cell. 153 (3): 590–600. doi:10.1016/j.cell.2013.03.025. PMC 3641580. PMID 23622243.
- ^ Supek F, Lehner B (July 2017). «Clustered Mutation Signatures Reveal that Error-Prone DNA Repair Targets Mutations to Active Genes». Cell. 170 (3): 534–547.e23. doi:10.1016/j.cell.2017.07.003. hdl:10230/35343. PMID 28753428.
- ^ Polak P, Lawrence MS, Haugen E, Stoletzki N, Stojanov P, Thurman RE, et al. (January 2014). «Reduced local mutation density in regulatory DNA of cancer genomes is linked to DNA repair». Nature Biotechnology. 32 (1): 71–75. doi:10.1038/nbt.2778. PMC 4116484. PMID 24336318.
- ^ Swenberg JA, Lu K, Moeller BC, Gao L, Upton PB, Nakamura J, Starr TB (March 2011). «Endogenous versus exogenous DNA adducts: their role in carcinogenesis, epidemiology, and risk assessment». Toxicol Sci. 120 Suppl 1 (Suppl 1): S130–45. doi:10.1093/toxsci/kfq371. PMC 3043087. PMID 21163908.
- ^ a b Hamilton ML, Guo Z, Fuller CD, Van Remmen H, Ward WF, Austad SN, Troyer DA, Thompson I, Richardson A (May 2001). «A reliable assessment of 8-oxo-2-deoxyguanosine levels in nuclear and mitochondrial DNA using the sodium iodide method to isolate DNA». Nucleic Acids Res. 29 (10): 2117–26. doi:10.1093/nar/29.10.2117. PMC 55450. PMID 11353081.
- ^ Ming X, Matter B, Song M, Veliath E, Shanley R, Jones R, Tretyakova N (March 2014). «Mapping structurally defined guanine oxidation products along DNA duplexes: influence of local sequence context and endogenous cytosine methylation». J Am Chem Soc. 136 (11): 4223–35. doi:10.1021/ja411636j. PMC 3985951. PMID 24571128.
- ^ a b c Zhou X, Zhuang Z, Wang W, He L, Wu H, Cao Y, Pan F, Zhao J, Hu Z, Sekhar C, Guo Z (September 2016). «OGG1 is essential in oxidative stress-induced DNA demethylation». Cell Signal. 28 (9): 1163–1171. doi:10.1016/j.cellsig.2016.05.021. PMID 27251462.
- ^ Poetsch AR (2020). «The genomics of oxidative DNA damage, repair, and resulting mutagenesis». Comput Struct Biotechnol J. 18: 207–219. doi:10.1016/j.csbj.2019.12.013. PMC 6974700. PMID 31993111.
- ^ D’Augustin O, Huet S, Campalans A, Radicella JP (November 2020). «Lost in the Crowd: How Does Human 8-Oxoguanine DNA Glycosylase 1 (OGG1) Find 8-Oxoguanine in the Genome?». Int J Mol Sci. 21 (21): 8360. doi:10.3390/ijms21218360. PMC 7664663. PMID 33171795.
- ^ Lan L, Nakajima S, Oohata Y, Takao M, Okano S, Masutani M, Wilson SH, Yasui A (September 2004). «In situ analysis of repair processes for oxidative DNA damage in mammalian cells». Proc Natl Acad Sci U S A. 101 (38): 13738–43. Bibcode:2004PNAS..10113738L. doi:10.1073/pnas.0406048101. PMC 518826. PMID 15365186.
- ^ Maeder ML, Angstman JF, Richardson ME, Linder SJ, Cascio VM, Tsai SQ, Ho QH, Sander JD, Reyon D, Bernstein BE, Costello JF, Wilkinson MF, Joung JK (December 2013). «Targeted DNA demethylation and activation of endogenous genes using programmable TALE-TET1 fusion proteins». Nat. Biotechnol. 31 (12): 1137–42. doi:10.1038/nbt.2726. PMC 3858462. PMID 24108092.
- ^ Ding N, Bonham EM, Hannon BE, Amick TR, Baylin SB, O’Hagan HM (June 2016). «Mismatch repair proteins recruit DNA methyltransferase 1 to sites of oxidative DNA damage». J Mol Cell Biol. 8 (3): 244–54. doi:10.1093/jmcb/mjv050. PMC 4937888. PMID 26186941.
- ^ a b Jiang Z, Lai Y, Beaver JM, Tsegay PS, Zhao ML, Horton JK, Zamora M, Rein HL, Miralles F, Shaver M, Hutcheson JD, Agoulnik I, Wilson SH, Liu Y (January 2020). «Oxidative DNA Damage Modulates DNA Methylation Pattern in Human Breast Cancer 1 (BRCA1) Gene via the Crosstalk between DNA Polymerase β and a de novo DNA Methyltransferase». Cells. 9 (1): 225. doi:10.3390/cells9010225. PMC 7016758. PMID 31963223.
- ^ Mortusewicz O, Schermelleh L, Walter J, Cardoso MC, Leonhardt H (June 2005). «Recruitment of DNA methyltransferase I to DNA repair sites». Proc Natl Acad Sci U S A. 102 (25): 8905–9. Bibcode:2005PNAS..102.8905M. doi:10.1073/pnas.0501034102. PMC 1157029. PMID 15956212.
- ^ a b Cuozzo C, Porcellini A, Angrisano T, Morano A, Lee B, Di Pardo A, Messina S, Iuliano R, Fusco A, Santillo MR, Muller MT, Chiariotti L, Gottesman ME, Avvedimento EV (July 2007). «DNA damage, homology-directed repair, and DNA methylation». PLOS Genet. 3 (7): e110. doi:10.1371/journal.pgen.0030110. PMC 1913100. PMID 17616978.
- ^ a b O’Hagan HM, Mohammad HP, Baylin SB (August 2008). «Double strand breaks can initiate gene silencing and SIRT1-dependent onset of DNA methylation in an exogenous promoter CpG island». PLOS Genet. 4 (8): e1000155. doi:10.1371/journal.pgen.1000155. PMC 2491723. PMID 18704159.
- ^ Ha K, Lee GE, Palii SS, Brown KD, Takeda Y, Liu K, Bhalla KN, Robertson KD (January 2011). «Rapid and transient recruitment of DNMT1 to DNA double-strand breaks is mediated by its interaction with multiple components of the DNA damage response machinery». Hum Mol Genet. 20 (1): 126–40. doi:10.1093/hmg/ddq451. PMC 3000680. PMID 20940144.
- ^ Russo G, Landi R, Pezone A, Morano A, Zuchegna C, Romano A, Muller MT, Gottesman ME, Porcellini A, Avvedimento EV (September 2016). «DNA damage and Repair Modify DNA methylation and Chromatin Domain of the Targeted Locus: Mechanism of allele methylation polymorphism». Sci Rep. 6: 33222. Bibcode:2016NatSR…633222R. doi:10.1038/srep33222. PMC 5024116. PMID 27629060.
- ^ Farris MH, Texter PA, Mora AA, Wiles MV, Mac Garrigle EF, Klaus SA, Rosfjord K (December 2020). «Detection of CRISPR-mediated genome modifications through altered methylation patterns of CpG islands». BMC Genomics. 21 (1): 856. doi:10.1186/s12864-020-07233-2. PMC 7709351. PMID 33267773.
- ^ Allen B, Pezone A, Porcellini A, Muller MT, Masternak MM (June 2017). «Non-homologous end joining induced alterations in DNA methylation: A source of permanent epigenetic change». Oncotarget. 8 (25): 40359–40372. doi:10.18632/oncotarget.16122. PMC 5522286. PMID 28423717.
- ^ Cromie GA, Connelly JC, Leach DR (December 2001). «Recombination at double-strand breaks and DNA ends: conserved mechanisms from phage to humans». Molecular Cell. 8 (6): 1163–74. doi:10.1016/S1097-2765(01)00419-1. PMID 11779493.
- ^ O’Brien PJ (February 2006). «Catalytic promiscuity and the divergent evolution of DNA repair enzymes». Chemical Reviews. 106 (2): 720–52. doi:10.1021/cr040481v. PMID 16464022.
- ^ Maresca B, Schwartz JH (January 2006). «Sudden origins: a general mechanism of evolution based on stress protein concentration and rapid environmental change». The Anatomical Record Part B: The New Anatomist. 289 (1): 38–46. doi:10.1002/ar.b.20089. PMID 16437551.
- ^ «CRISPR gene-editing tool has scientists thrilled – but nervous» CBC news. Author Kelly Crowe. 30 November 2015.
External links[edit]
![]()
This audio file was created from a revision of this article dated 17 June 2005, and does not reflect subsequent edits.
- Media related to DNA repair at Wikimedia Commons
- Roswell Park Cancer Institute DNA Repair Lectures
- A comprehensive list of Human DNA Repair Genes
- 3D structures of some DNA repair enzymes
- Human DNA repair diseases
- DNA repair special interest group
- DNA Repair Archived 12 February 2018 at the Wayback Machine
- DNA Damage and DNA Repair
- Segmental Progeria
- DNA-damage repair; the good, the bad, and the ugly
Механизмы исправления ошибок во время репликации ДНК и ее репарация вследствие повреждений на протяжении всего жизненного цикла клетки.
Основные моменты:
-
Клетки имеют различные механизмы предотвращения возникновения мутаций – необратимых изменений в ДНК
-
В процессе синтеза ДНК, большинство ДНК-полимераз «проверяют свою работу» и проводят замену бо́льшей части ошибочно вставленных нуклеотидов. Этот процесс можно назвать исправлением ошибок.
-
Сразу после синтеза ДНК любые оставшиеся ошибочные нуклеотиды обнаруживаются и заменяются в так называемом процессе репарации ошибочно спаренных нуклеотидов.
-
Если ДНК повреждена, она может быть восстановлена с помощью различных механизмов, например, путём прямой репарации, эксцизионной репарации или путём восстановления двухцепочечных разрывов
- пострепликативной репарации.
Введение
Как ДНК связана с раком? Рак возникает при неконтролируемом делении клеток, когда игнорируются клеточные «стоп»-сигналы, что приводит к образованию опухоли. Это неправильное поведение клеток вызвано накопившимися мутациями — необратимыми изменениями последовательности ДНК клетки.
На самом деле, ошибки в процессе репликации и повреждения ДНК возникают в клетках нашего тела постоянно. Однако в большинстве случаев они не приводят к раку и даже не вызывают мутаций, такие ошибки обычно обнаруживаются и исправляются в процессе репарации ДНК. Если же повреждение исправить не удаётся, то в клетке включается механизм самоуничтожения — (апоптоз), который предотвращает передачу поврежденной ДНК дочерним клеткам.
Мутации возникают и передаются дочерним клеткам только тогда, когда эти механизмы не справляются. В частности, рак возникает в случае накопившихся в одной клетке мутаций генов, связанных с делением.
В этой статье мы подробно рассмотрим механизмы, используемые клетками для исправления ошибок, которые возникают в процессе репликации. К ним относятся:
-
Исправление ошибок – процесс, который возникает во время репликации ДНК.
-
Репарация ошибочно спаренных нуклеотидов, которая происходит сразу же после репликации ДНК.
-
Механизмы репарации, которые выявляют и исправляют повреждения ДНК на протяжении всего клеточного цикла
Исправление ошибок
ДНК-полимеразы — это ферменты, участвующие в репликации ДНК. Во время копирования ДНК большинство ДНК-полимераз «проверяют», корректный ли нуклеотид они добавляют. Этот процесс называется исправлением ошибок. Если полимераза обнаружит, что был добавлен неправильный нуклеотид, она сразу же удалит и заменит его и только после этого продолжит синтез ДНКstart superscript, 1, end superscript.
Репарация ошибочно спаренных нуклеотидов
Процесс исправления избавляет от основной массы ошибок, но не от всех. После создания новой ДНК запускается механизм репарации ошибочно спаренных нуклеотидов — удаления и замены ошибочно спаренных нуклеотидов, оставшихся в результате репликации. Исправление несоответствий между парами оснований также может включать в себя исправление небольших вставок и делеций, возникающих вследствие «соскальзывания» полимеразы с исходной цепи squared.
Как происходит восстановление неправильно спаренных нуклеотидов? Во-первых, белковый комплекс распознаёт неправильно спаренный нуклеотид и связывается с ним. Другой комплекс разрезает ДНК в области несовпадения, а ещё одна группа ферментов отщепляет некорректный нуклеотид вместе с небольшим участком вокруг него. Затем ДНК-полимераза заполняет этот пробел правильными нуклеотидами, а фермент ДНК-лигаза сшивает разрывы в цепиsquared.
Удивительно: как белки, участвующие в восстановлении ДНК, определяют, «кто прав» во время репарации ошибочно спаренных нуклеотидов? То есть, когда два основания неправильно соединены (как G (гуанин) и T (тимин) на рисунке выше), какое из этих двух оснований должно быть удалено и заменено?
У бактерий можно отличить исходную и дочернюю цепи ДНК по метилированным основаниям. На исходной цепи ДНК есть метильные (minus, start text, C, H, end text, start subscript, 3, end subscript) группы, присоединенные к некоторым из ее оснований, а у дочерней цепи таких групп еще нетcubed.
У эукариот процессы, позволяющие идентифицировать исходную цепь при устранении несоответствий, включают распознавание одноцепочечных разрывов, которые обнаруживаются только у дочерней цепи cubed.
Механизмы репарации ДНК
С ДНК может что-нибудь случиться практически в любой момент жизни клетки, а не только во время репликации. Фактически, ДНК постоянно повреждается из-за воздействия внешних факторов: ультрафиолетового излучения и радиации, химических веществ, не говоря уже о спонтанных процессах, которые протекают даже без вмешательства окружающей среды!start superscript, 4, end superscript
К счастью, наши клетки имеют механизмы восстановления, с помощью которых они находят и исправляют большинство повреждений ДНК. Можно выделить несколько типов репарации:
-
Прямая репарация. Некоторые повреждения ДНК, вызванные химическими реакциями, могут быть «исправлены» находящимися в клетке ферментами.
-
Эксцизионная репарация. Повреждение одного или нескольких нуклеотидов ДНК часто исправляется удалением и заменой поврежденного участка. При эксцизионной репарации оснований удаляется только поврежденное основание. В случае эксцизионной репарации нуклеотидов, как и в случае репарации ошибочно спаренных нуклеотидов, которое мы рассмотрели выше, удаляются целиком нуклеотиды.
-
Репарация двухцепочечных разрывов: Существуют два основных способа: негомологичное соединение концов и гомологичная рекомбинация. Они используются для восстановления двухцепочечных разрывов ДНК (когда вся хромосома разделяется на две части).
Прямая репарация
В некоторых случаях клетка может исправить повреждение ДНК, обратив вызвавшую его реакцию. Дело в том, что «повреждение ДНК» — это, как правило, присоединение к ней лишней группы в результате химической реакции.
Например, гуанин (G) может подвергаться реакции с присоединением метильной (minus, start text, C, H, end text, start subscript, 3, end subscript) группы к атому кислорода в азотистом основании. Если это не исправить, метил-содержащий гуанин будет связываться с тимином (Т), а не с цитозином (С) во время репликации ДНК. К счастью, у людей и многих других организмов есть фермент, который может удалить метильную группу, обратив реакцию, и тем самым вернуть азотистое основание в нормальное состояниеstart superscript, 5, end superscript.
Эксцизионная репарация оснований
Эксцизионная репарация оснований — это механизм, используемый для обнаружения и удаления определенных типов поврежденных азотистых оснований. Ключевую роль в нем играет группа ферментов, называемых гликозилазами. Каждая гликозилаза обнаруживает и удаляет определенный вид поврежденных оснований.
Например, в процессе реакции дезаминирования цитозин может превратиться в урацил — основание, обычно встречающееся только в РНК. Во время репликации ДНК урацил будет соединяться с аденином, а не с гуанином (в отличие от цитозина), поэтому такое превращение может привести к возникновению мутацииstart superscript, 5, end superscript.
Для предотвращения подобных изменений гликозилаза, являющаяся частью сигнального пути эксцизионной репарации, обнаруживает и удаляет дезаминированные цитозины. После того, как основание было удалено, удаляется и оставшаяся часть нуклеотида, а другие ферменты заполняют пробелstart superscript, 6, end superscript.
Эксцизионная репарация нуклеотидов
Эксцизионная репарация нуклеотидов — это еще один способ удаления и замены поврежденных оснований. В результате нее обнаруживаются и корректируются повреждения, которые искажают форму двойной спирали ДНК. Например, азотистые основания могут измениться, присоединив к себе громоздкие группы атомов, в частности, в результате воздействия химических веществ, содержащихся в сигаретном дымеstart superscript, 7, end superscript.
Эксцизионная репарация нуклеотидов также используется для устранения повреждений, вызванных ультрафиолетовым излучением, например, при получении солнечного ожога. Под воздействием УФ-излучения цитозин и тимин могут вступать в реакцию с соседними основаниями, которые также являются цитозином или тимином, образуя при этом связи, изменяющие форму двойной спирали и вызывающие ошибки в процессе репликации ДНК. Наиболее распространенный тип таких связей — тиминовый димер — он состоит из двух тиминовых оснований, вступающих в реакцию друг с другом и образующих химическую связьstart superscript, 8, end superscript.
При эксцизионной репарации нуклеотидов поврежденные нуклеотиды удаляются вместе с соседними нуклеотидами. В этом процессе хеликаза (фермент, раскручивающий ДНК) раскрывает ДНК, образуя пузырь, а ферменты, разрезающие ДНК, отсекают поврежденную часть пузыря. Полимераза заполняет пробел, а лигаза сшивает разрыв в цепиstart superscript, 9, end superscript.
Репарация двухцепочечных разрывов
Некоторые факторы окружающей среды, например, радиация, могут вызывать разрывы обеих цепочек ДНК (разделение хромосомы на две части). Такие повреждения ДНК, если верить комиксам, ведут к появлению супергероев, но могут встречаться и после реальных катастроф, например, Чернобыльской.
Двухцепочечные разрывы опасны, потому что большие сегменты хромосом и сотни содержащихся в них генов могут быть потеряны, если разрыв не будет восстановлен. Существует два способа восстановления двухцепочечных разрывов ДНК: негомологичное соединение концов и гомологичная рекомбинация.
При негомологичном соединении концов два разорванных конца хромосомы просто склеиваются обратно. Этот механизм восстановления является «грубым» и неточным, в результате в месте разрыва, как правило, либо теряются нуклеотиды, либо добавляются лишние, что может привести к мутациям. Но это в любом случае лучше потери целого фрагмента хромосомыstart superscript, 10, end superscript.
При гомологичной рекомбинации для восстановления разрыва используется фрагмент из гомологичной хромосомы, который соответствует поврежденной хромосоме (или из сестринской хроматиды, если ДНК была реплицирована). В этом процессе две хромосомы объединяются, и неповрежденная область гомологичной хромосомы или хроматиды используется в качестве матрицы для замены поврежденной области. Гомологичная рекомбинация работает «чище», точнее, чем негомологичное соединение концов, и обычно не приводит к образованию мутацийstart superscript, 11, end superscript.
Репарация ДНК и заболевания человека
Доказательства важности механизмов репарации получены на основе генетических заболеваний человека. Во многих случаях мутации в генах, которые кодируют белки, участвующие в репарации, связаны с наследственным раком. Например:
-
Наследственный неполипозный колоректальный рак (также называемый синдромом Линча) вызван мутациями в генах, кодирующих белки, которые участвуют в репарации ошибочно спаренных нуклеотидовstart superscript, 12, comma, 13, end superscript. Поскольку такие нуклеотиды не восстанавливаются, у людей, страдающих этим синдромом, мутации накапливаются гораздо быстрее, чем у здоровых. Это может привести к развитию опухолей толстой кишки.
-
Люди с пигментной ксеродермой очень чувствительны к ультрафиолетовому излучению. Это вызвано мутациями в белках, участвующих в эксцизионной репарации нуклеотидов. Когда они не функционируют, димеры тимина и другие виды повреждений, вызванные ультрафиолетовым излучением, перестают восстанавливаться. У людей с пигментной ксеродермой после нескольких минут пребывания на солнце могут возникнуть сильные солнечные ожоги, и около половины из них заболевают раком кожи в возрасте до 10 лет, если только они не избегают солнечных лучейstart superscript, 14, end superscript.
рис.1 Модифицированные азотистые основания ДНК, удаляемые ДНК–гликозилазами: а – урацил; б – гипоксантин; в – 5–гидроксицитозин; г – 2,5-диамино-4-формамидопиримидин; д – 7,8-дигидро-8-оксогуанин; е – мочевина; ж – тимингликоль; з – 5-формилурацил; и – 5-гидроксиметилурацил; к – 3-метиладенин; л – 7-метилгуанин; м – 2-метилцитозин [Партушев, 2000].
АP-эндонуклеаза создает ник (одноцепочечный разрав) с 3’-ОН и 5’-dRP концами. Для большинства млекопитающих характерен
тип BER репарации с включением одного нуклеотида. В этом случае ДНК-полимераза β вставляет 1 нуклеотид в 3’-конец праймера и затем удаляет 5’-dRP фрагмент с помощью своей dRP-лиазной активности. Получившийся ник сшивается ДНК-лигазой. Альтернативным является путь так называемой длинно-заплаточной BER репарации. Он реализуется в тех случаях, когда 5’-dRP фрагмент модифицирован и не может быть удален с помощью ДНК–полимеразы β. В этом случае Pol β проводит синтез ДНК с вытеснением запирающей цепи, 2 — 13 нуклеотидов. Образующийся в результате свисающий 5’-конец ДНК выщепляется флэпэндонуклеазой FEN1. Pol бета, Pol дельтаи Pol епсилон могут вести ситнез ДНК во время длинно-заплаточной репарации. Идентичность полимераз, вовлеченных в этот процесс in vitro, еще не ясна, но показано, что Pol β всегда инициирует синтез ДНК.
Зависимость того, какой из путей BER реализуется, определяется бифункциональными ДНК-гликозилазами, которые имеют дополнительную
АP-лиазную активность. При комбинации ДНК-гликозилазной, АP-лиазной и АP-эндонуклеазной активностей внутри одного фермента в поврежденной цепи ДНК образуется однонуклеотидная брешь с 3’-ОН и 5’-P концами, которую может застроить Pol β.
Дополнительная сложность в понимании механизмов переключения внутри BER состоит в том, что две новые полимеразы Pol i и Pol лямбда также имеют dRP-лиазнуюактивность. Таким образом, можно предположить, что они, как и Pol бета, являются участниками тех процессов репарации, где необходимо выщепление dRP фрагмента. Pol лямбда является близким гомологом Pol бета со сходными ферментативными свойствами. Также как и Pol бета, она лишена 3’—>5’экзонуклеазной активности и имеет низкую процессивность синтеза на частичном ДНК-дуплексе, содержащем свисающий 5’-участок матрицы, процессивный синтез возможен в брешах с 5’-P концами. Таким образом, Pol лямбда является подходящим кандидатом для BER синтеза. Более того,
Pol лямбда заменяет Pol бета в реконструированных BER системах, репарирующих урацил-содержащие ДНК, iv vitro. Однако, клетки мышей Pol лямбда -/- не чувствительны к обработке перекисью или метилметансульфонатом, агентам, которые, помимо других типов повреждений, продуцируют АР-сайты и окисленные формы оснований. Отсюда можно заключить, что Pol лямбда не является необходимой для BER в клетках, где есть, Pol дельта и Pol епсилон. Интересно, что Pol лямбда может эффективно процессировать ДНК при очень низкой концентрации dNTP (около 1 мкМ), что может означать ее участие в любых клеточных процессах в фазе G0 при низкой концентрации dNTP. В поддержку этой гипотезы выступает тот факт, что экспрессия Pol лямбда зависит от клеточного цикла, и наибольшее количества белка экспрессируется при переходе из S- в М-фазу и в спокойных клетках.
Pol i принадлежит к Y семейству полимераз. Основная функция полимераз этого семейства – утилизация ДНК-повреждений, блокирующих работу репликативных ДНК-полимераз. Однако,
исходя из некоторых свойств Pol i, можно предположить, что этот фермент является участником альтернативного процесса BER репарации. Pol i обладает низкой поцессивностью, лишена 3’>5’экзонуклеазной активности и способна застраивать брешь в 1 — 5 нуклеотидов iv vitro. Pol i может замещать Pol бета в реконструированных системах BER, репарирующих урацил-содержащие ДНК, iv vitro, благодаря наличию dRP-лиазной и ДНК-полимеразной активностей. Кинетические исследования реакции нуклеотидного встраивания овыявили дополнительные данные, свидетельствующие в пользу того, что Pol i может принимать участие в репарации. Pol i вставляет dTMP напротив А в ДНК-матрицу с большей эффективностью, чем любой другой дезоксинуклеозидмонофосфат, при этом точность ДНК-синтеза сопоставима с параметрами для Pol бета. Основываясь на этих данных, можно предположить, что Pol i – участник BER репарации оснований уридина, который образуется после встраивания dUMP напротив А во время ошибочной репликации. Pol i также
может вставлять dGМP напротив Т со скоростями, близкими к скорости включения корректного нуклеотида. Более того, на матрице, содержащей 2 или более последовательных Т, вторым встраиваемым нуклеотидом оказался dGMP. Подобные результаты служат основой для гипотезы, что Pol i может быть участником альтернативной BER репарации в тех слсучаях, когда dG был ошибочно удален гликозилазой из G-T или G-U некомплементарной пары, образовавшейся в результате дезаминации 5-метилцитозина или цитозина. Возможная роль Pol i в трансляции синтеза ДНК и процессе соматических гипермутаций рассмотрена ниже.
Возможность участия пяти ядерных полимераз в BER, три из которых обладают dRP-лиазной активностью, была мало изучена в клетках и на модельных животных, лишенных более чем одной полимеразы. Поэтому данные о субстратной специфичности и взаимодействии полимеразо-акцепторных белков, необходимых в BER, недостаточны. Помимо BER репарации, Pol бетта принимает участие в репарации одноцепочечных разрывов; функционирование
этого процесса нарушено у пациентов с наследственной спинномозговой атаксией. Одноцепочечные разрывы генерируются эндогенными или экзогенными агентами. Такие разрывы часто содержат однонуклеотидные бреши с 3’ и/или 5’ модифицированными концами, то есть очень похожи на субстраты BER. Так вот, интересно посмотреть, способны ли другие полимеразы с подобным набором исходных данных (Pol i и Pol лямбда) принимать участие в репарации одноцепочечных разрывов.
Четвертой ДНК-полимеразой в клетках человека, обладающей dRP-лиазной активностью, является митохондриальная ДНК-полимераза гамма. Pol гамма – единственная ДНК-полимераза, обнаруженная в митохондриях, следовательно, она ответственна за все преобразованиях ДНК, происходящие в этой органелле. Митохондрия является объектом интенсивного повреждения ДНК активными формами кислорода, генерируемыми во время окислительного фосфорилирования. Эти повреждения эффективно репарируются набором митохондриальных белков, которые включают в себя АP-эндонуклеазу, Pol гамма,
мтДНК-лигазу. Сам процесс репарации сходен с однонуклеотидным процессом BER в ядерной ДНК.
Таблица. ДНК-гликозилазы и эндонуклеазы клеток микроорганизмов и человека, участвующие в BER.
В универсальном механизме эксцизионной репарации как прокариоты, так и эукариоты гидролизуют 3–5-ю фосфодиэфирную связь с 3′-конца отповреждения. При этом прокариоты гидролизуют также 8-ю связьот 5’-конца измененного нуклеотида, тогда как у эукариотических организмовпроисходит одноцепочечный разрыв на расстоянии 21–25 нуклеотидов отповреждения со стороны его 5’-конца. Таким образом, прокариоты удаляютизмененный нуклеотид в составе 12–13-членных олигомеров, тогда как
эукариоты – в составе одноцепочечных фрагментов ДНК длиной в 27–29 нуклеотидов. Ферментная система, вносящая такие двойные одноцепочечные
разрывы, получила название эксцизионной нуклеазы (эксцинуклеазы). Образующаяся в молекуле репарируемой ДНК одноцепочечная брешь далее заполняется с помощью ДНК-полимеразы, а фосфодиэфирная связь в остающемся одноцепочечном разрыве восстанавливается ДНК-лигазой.
NER устраняет ДНК-повреждения посредством вырезания олигонуклеотида, содержащего это повреждение, с образованием бреши в ДНК размером ~ 30 нуклеотидов. Эта брешь застраивается одной или двумя полимеразами В семейства Pol дельта или Pol епсилон. Любая из этих ДНК-полимераз способна застраивать подобную брешь в реконструированной системе, содержащей очищенные белки млекопитающих, in vitro. При изучении NER в системе фибробластов человека с нарушенной проницаемостью и в ядерном экстракте HeLa клеток обнаружили, что обе полимеразы необходимы для восстановления ДНК после УФ-облучения. Исследования
в дрожжевых системах показали, что та или другая ДНК-полимераза существенно дополняют репарацию УФ-поврежденной ДНК. Более поздние исследования показали, что оба фермента необходимы для эффективной NER репарации в дрожжевых экстрактах. Pol дельта и Pol епсилон способны вести процессивный синтез ДНК при наличии дополнительного фактора PCNA, который «надевается» на ДНК с помощью пятисубъединичного комплекса RFC. Репарационный синтез как Pol дельта, так и Pol епсилон также требует присутствия этих факторов процессивности.
Эксцизионная репарация нуклеотидов: введение
ЭР — эксцизионная репарация ДНК; ЭР-комплекс включает ПАРП , XRCC1 , ДНК-лигазу III и ДНК-полимеразу бета .
Два пути осуществления NER:
1. Гидролиз фосфодиэфирной связи по 3′- или 5′- концу на некотором расстоянии от ошибочно спаренного (поврежденного) нуклеотида, который далее целиком удаляется под действием 5′->3′- (или 3′->5′-) экзонуклеазы, гидролизующей
цепь ДНК нуклеотид за нуклеотидом в соответствующем направлении от первоначального одноцепочечного разрыва в репарируемой ДНК. Образующаяся брешь далее заполняется ДНК-полимеразой. Такой механизм репарации реализуется у E. coli и человека для вырезания неповрежденных (немодифицированных) ошибочно спаренных нуклеотидов. Механизм последовательного эндо- и экзонуклеазного расщепления ДНК не используется для удаления поврежденных (измененных) нуклеотидов. Это связано с тем, что такие нуклеотиды часто являются ингибиторами экзонуклеаз.
2. Функционирует у всех исследованных видов организмов и заключается в использовании ферментной системы, которая вносит одноцепочечные разрывы по обе стороны от поврежденного нуклеотида на некотором расстоянии от него с последующим удалением одноцепочечного фрагмента ДНК, содержащего измененный нуклеотид.
Гидролизуется 3-5-фосфодиэфирную связь с 3′-конца от повреждения. При этом прокариоты гидролизуют также 8-связь от 5′-конца
измененного нуклеотида, тогда как у эукариотических организмов происходит одноцепочечный разрыв на расстоянии 21-25 нуклеотидов от повреждения со стороны его 5′-конца. Таким образом, прокариоты удаляют измененный нуклеотид в составе 12-13-членных олигомеров, тогда как эукариоты — в составе одноцепочечных фрагментов ДНК длиной в 27-29 нуклеотидов. Ферментная система, вносящая такие двойные одноцепочечные разрывы, получила название эксцизионной нуклеазы (эксцинуклеазы) . Образующаяся в молекуле репарируемой ДНК одноцепочечная брешь далее заполняется с помощью ДНК-полимеразы, а фосфодиэфирная связь в остающемся одноцепочечном разрыве восстанавливается ДНК-лигазой.
В отличие от ЭРО и прямых реверсий, которые специфичны к достаточно узкому кругу повреждений ДНК, система ЭРН, хотя и с разной эффективностью, удаляет все возможные повреждения, и потому роль ее в поддержании стабильности генома велика. ЭРН детально изучена у E. coli и активно изучается в клетках
дрожжей и человека, причем в последних благодаря раскрытию генетической природы таких заболеваний, как пигментная ксеродерма ( ХР ), синдром Кокейна ( СS ) и заболевания трихотиодистрофия ( ТТD)
В ЭРН задействовано 6-8 генов у E. coli и до 30 у человека.
Если повреждение в транскрибируемой (матричной) нити ДНК задерживает продвижение РНК-полимеразы, то специальный белковый фактор TRCF ( transcription-repair coupling factor ) сталкивает РНК-полимеразу и связывает с этим местом комплекс ферментов репарации. См. Сопряжение транскрипции и репарации ДНК у E/coli.
Ферментный ансамбль осуществляющий первые 3 стадии процесса NER называется эксцинуклеазой.
В системе эксцизионной репарации ДНК путем удаления нуклеотидов поврежденные азотистые основания вырезаются в составе олигонуклеотидов.
NER: регуляция
Для клеток животных не характерен SOS-ответ, свойственный клеткам E. coli и представляющий собой суммарную реакцию бактериальной клетки на повреждение ДНК различными агентами,
проявляющийся в усилении транскрипции генов NER. Посттрансляционные модификации белков репарации, происходящие в ответ на повреждение ДНК, не влияют на активность эксцинуклеазы человека.
Обнаружено, что повреждения ДНК стабилизирует белок р53- белок-супрессор опухолевого роста, являющийся регулятором транскрипции. Имеются данные о том, что белок р53 может взаимодействовать с белками XPB и RPA, необходимыми для NER. Однако клетки с инактивированными генами р53 (p53(-/-)), как и клетки дикого типа, эффективно удаляют из поврежденной ДНК два основных фотопродукта, возникающих под действием УФ-света, и обладают такой же устойчивостью к УФ. Поэтому считается, что белок р53 не оказывает прямого влияния на NER. Белки Cdk7 и циклин H, которые образуют Cdk-активирующую киназу, входят в состав комплекса TFIIH, что позволяет предполагать наличие связи репарации ДНК с фазами клеточного цикла.
ЭРН (NER): сопряжение с транскрипцией (ТЭРН)
Транскрибируемые последовательности нуклеотидов ДНК, особенно
в матричной цепи, репарируются с большей скоростью, чем нетранскрибируемые последовательности. В клетках больных с синдромом Кокайна не наблюдается такой асимметрии в репарации.
В клетках E. coli белковый фактор, кодируемый геном mfd и сопрягающий транскрипцию и репарацию, замещает остановившиеся перед повреждением молекулы РНК-полимеразы, что приводит к диссоциации транскрипционного комплекса. При этом он привлекает экзонуклеазный репаративный комплекс к поврежденному участку ДНК. См. Сопряжение транскрипции и репарации ДНК (ТЭРН) у E/coli
В клетках животных ген CSB кодирует белок с молекулярной массой 160 кДа, который содержит хеликазный домен и, возможно, выполняет те же функции, что и белок Mfd у E. coli . На основе поведения клеток с мутантными генами белков CSA и CSB разработана модель механизма, с помощью которого обеспечивается асимметричная репарация цепей ДНК. В соответствии с этой моделью РНК-полимераза II , остановившаяся в процессе транскрипции
перед поврежденным участком ДНК, распознается комплексом CSA-CSB и перемещается в сторону от повреждения без разрушения четвертичной структуры транскрипционного комплекса. Одновременно комплекс CSA-CSB привлекает компоненты репаративной системы XPA и TFIIH к месту повреждения ДНК и помогает сборке эксцинуклеазного комплекса. Нуклеотиды поврежденной цепи вырезаются, и брешь репарируется. После этого РНК-полимераза в составе транскрипционного комплекса продолжает транскрипцию.
NER: механизм
Процесс NER можно разделить на четыре этапа: а) распознавание поврежденного участка ДНК; б) двойное надрезание (инцизия) цепи ДНК по обеим сторонам поврежденного участка и его удаление (эксцизия); в) заполнение бреши в процессе репаративного синтеза; г) лигирование оставшегося одноцепочечного разрыва ДНК.
NER человека распознает и удаляет одиночные ошибочно спаренные нуклеотиды, а также петли длиной в 1-3 нуклеотида. NER человека способна различать цепи ДНК в случае распознавания поврежденных
нуклеотидов. Показано, что при наличии в ДНК димеров тимина циклобутанового типа вырезание нуклеотидов происходит исключительно из поврежденной цепи. Механизм такого распознавания в настоящее время неизвестен, как и молекулярный механизм узнавания самих поврежденных оснований. Система способна распознавать повреждения как сильно, так и слабо деформирующие вторичную структуру ДНК. При этом не обнаружена линейная зависимость между коэффициентом специфичности нуклеазы (kcat/km) и уровнем деформации двойной спирали ДНК. Показано, что в процессе распознавания участвуют белковые комплексы XPA/RPA, которые преимущественно связываются с поврежденной ДНК, и TFIIH, обладающий АTP-зависимой ДНК-расплетающей активностью. Последний взаимодействует с поврежденным участком ДНК и по аналогии с соответствующим механизмом у E. coli локально раскручивает ДНК, создавая основной преинцизионный комплекс с поврежденной ДНК.
Установлено, что три фермента репарации, обладающие узкой
субстратной специфичностью: ДНК-фотолиаза (удаление пиримидиновых димеров), урацилгликозилаза (удаление урацила из ДНК) и экзонуклеаза III (гидролиз ДНК в AP-сайтах), втягивают поврежденный участок из двойной спирали в полость фермента, что приводит кофактор или аминокислотные остатки активного центра этих ферментов в непосредственный контакт с расщепляемыми связями ДНК. Не исключено, что система эксцинуклеазы действует таким же образом.
Основные этапы функционирования NER, следующие за распознаванием поврежденного участка ДНК, представлены на рис. I.59. После связывания комплекса XPA-RPA с измененным участком ДНК, XPA взаимодействует с комплексом TFIIH, который создает преинцизионный комплекс, что сопряжено с гидролизом ATP. ATP-зависимое расплетание ДНК комплексом TFIIH подготавливает ее к взаимодействию с двумя XP-белками, обладающими нуклеазной активностью. XPG связывается с TFIIH и вносит одноцепочечный разрыв с 3′-конца повреждения. Аналогично комплекс ERCC1-XPF
взаимодействует с XPA в составе преинцизионного комплекса и способствует одноцепочечному разрыву с 5′-конца повреждения. Образование обоих разрывов является ATP-зависимым, и их расположение на ДНК высокоспецифично. Как правило, происходят разрывы 5-й и 24-й фосфодиэфирных связей соответственно от 3′- и 5′-концов поврежденных участков. Однако расположение точек разрывов может варьироваться.
Таким образом, в результате подобных одноцепочечных надрезов ДНК может освобождаться фрагмент длиной 24-32 нуклеотида с преобладанием фрагментов длиной 27-29 нуклеотидов. На расположение сайтов одноцепочечных разрывов влияют характер повреждения и последовательности нуклеотидов (контекст), окружающих поврежденный участок. Ту же самую картину инцизии обнаруживают in vivo в ооцитах Xenopus и у Schizosaccharomyces pombe. На этом основании делают вывод об универсальном механизме эксцизионной репарации у эукариот.
Репаративный синтез ДНК у человека является PCNA-зависимым,
т.е. может осуществляться с участием ДНК-полимераз Polдельта и Polэпсилон. PCNA связывается с системой праймер-матрица под действием фактора репликации RFC, откуда следует, что последний также участвует в репаративном синтезе ДНК. В опытах с бесклеточными системами моноклональные антитела к Polдельта специфически подавляют репаративный синтез. Однако оказалось, что в тех же высокоочищенных бесклеточных системах вместо Polдельта с аналогичным эффектом могут быть использованы Polэпсилон и даже фрагмент Кленова ДНК-полимеразы I E. coli. Это означает, что реконструированные из очищенных компонентов бесклеточные системы лишь в ограниченной степени имитируют биохимические процессы, происходящие в живых клетках. Считается, что обе ДНК-полимеразы — Polдельта и Polэпсилон участвуют в репаративном синтезе ДНК у человека.
ПАРП: модели управления процессом эксцизионной репарации ДНК
ПАРП — один из первых ядерных факторов, распознающих повреждение ДНК , и поэтому в идеальном случае управляет запуском механизма
репарации ДНК в живых клетках с места повреждения ДНК. Эта модель поддерживается идентификацией ЭР -комплекса, включающего ПАРП , XRCC1 , ДНК-лигазу III и ДНК-полимеразу бета . Присутствие ПАРП в таком мультипротеиновом комплексе доказывает, что этот фермент может направлять аппарат репарации ДНК к сайтам повреждения ДНК in vivo и облегчает осуществление репарации по этому пути.
XRCC1-белок действует как молекулярные «леса», формируя ЭР-комплекс путем индивидуального взаимодействия с каждым компонентом. С тех пор, как была обнаружена возможность поли(АДФ-рибозил)ирования XRCC1-белка in vitro, можно предположить, что ПАРП способен регулировать активность комплекса путем модифицирования XRCC1-белка in vivo и нарушать его способность взаимодействовать с другими компонентами комплекса. Было показано, что сверхэкспрессия XRCC1-белка подавляет активность ПАРП в живых клетках.
Аналогично ДНК-лигаза III ингибирует активность ПАРП in vitro, когда ее количества превышают количества ПАРП.
ПАРП может также рекрутировать факторы репарации ДНК путем модификации хроматиновых белков. Длинные цепи ПАР действительно способны направить ферменты репарации к сайтам разрывов ДНК значительно быстрее, нежели если они ищут повреждение сами по всему ядру. Такая модель согласуется с активностью ПАРП перед и/или после удаления поврежденных оснований.
NER: структура и функции белков
В табл. I.21 суммированы свойства белков животных, участвующих в NER. Большинство этих белков существует in vivo в виде комплексов, поэтому ферментативные активности, обнаруживаемые у отдельных белков в очищенном состоянии, могут не иметь прямого отношения к их функциям в системе NER.
XPA — белок с молекулярной массой 31 кДа, обладает доменом типа «цинковые пальцы», участвует в распознавании поврежденного участка ДНК, взаимодействует с другими компонентами системы и может функционировать в качестве фактора нуклеации для экзонуклеазы. XPA взаимодействует своим N-концевым доменом
с гетеродимером ERCC1-XPF, а С-концевым доменом — с TFIIH.
Белок RPA (HSSB) образует комплекс с XPA и усиливает его специфичность в отношении поврежденной ДНК. RPA (HSSB) — тример, состоящий из белковых субъединиц р70, р34 и р11, необходим для репликации ДНК и репаративного синтеза, а также для прохождения этапа двойного надреза ДНК во время эксцизионной репарации. Он обладает умеренным сродством к поврежденной ДНК.
TFIIH — олигомерный комплекс, в состав которого входят белки р89, р80, р62, р44, р41, р38 и р34. Этот белковый комплекс был открыт как один из семи основных факторов транскрипции, необходимых для эффективного функционирования РНК-полимеразы II . Субъединица р89 идентична белку репаративного комплекса XPB. Обнаружено отсутствие функциональной комплементации между бесклеточными экстрактами клеток с мутантными белками XPB и XPD, определяемой по восстановлению репарирующей активности в смешанных экстрактах. Комплекс TFIIH представляет собой
фактор репаративной системы. Белки XPB и XPD являются ДНК-зависимыми АТРазами, обладают хеликазными доменами и могут (как и сам фактор TFIIH) вызывать диссоциацию гибридов, образованных между короткими фрагментами ДНК и одноцепочечной ДНК.
XPC — белок с молекулярной массой около 125 кДа, существует в виде гетеродимера в комплексе с белком р58, который является гомологом белка Rad23 дрожжей (HHR23B). XPC слабо связывается с TFIIH и очень прочно — с одноцепочечной ДНК.
ERCC1/XPF — прочный белковый комплекс, с которым взаимодействует белок XPA, обладающий эндонуклеазной активностью, специфичной в отношении одноцепочечной ДНК.
XPG — белковый комплекс, обладающий эндонуклеазной активностью, специфичной в отношении одноцепочечной ДНК; вовлекается в эксцизионный комплекс посредством взаимодействия с TFIIH и RPA.
ЭРН (NER): генетика
Гены NER E. coli, uvrA, uvrB и uvrC не обнаруживают гомологии с соответствующими генами человека. Гены NER дрожжей и человека
высокогомологичны, и энзимология эксцизионной репарации также обладает большим сходством. По крайней мере, три заболевания у человека вызываются генетическими нарушениями системы эксцизионной репарации: пигментная ксеродерма, синдром Кокейна и трихотиодистрофия.
Кожа больных пигментной ксеродермой обладает повышенной чувствительностью к дневному свету, что проявляется в виде фотодерматозов, включая рак кожи. В ряде случаев отмечены аномалии нервной системы, причиной которых являются мутации в одном из семи генов: XPA, XPB, XPC, XPD, XPE, XPF, XPG. Однако описаны больные с классическими симптомами пигментной ксеродермы, но с ненарушенной системой NER. Для клеток этих больных характерны изменения в так называемой пострепликативной репарации .
Больным с синдромом Кокейна присущи нарушения роста, умственная отсталость, катаракты, повышенная чувствительность к свету с сопутствующими дерматозами. Обнаружены мутации в двух группах генов, приводящие к этому заболеванию. У больных
с мутантными генами CS-A или CS-B клетки способны нормально репарировать УФ-повреждения ДНК. У другой группы больных обнаружены мутации в генах XPB, XPD или XPG.
У больных трихотиодистрофией со смешанными симптомами выявлены мутации в генах XPB или XPD. Классические симптомы этого заболевания, по-видимому, являются следствием мутации в гене транскрипционного фактора TFIIH.
Получение мутантов с измененной NER у грызунов позволило разбить такие гены на 11 групп комплементации, большинство из которых соответствует группам комплементации XP и CS человека. Часть соответствующих генов человека удалось клонировать, используя их способность исправлять (комплементировать) генетические дефекты в культивируемых мутантных клетках грызунов. Эти гены получили название кросс-комплементирующих генов эксцизионной репарации ( ERCC — excision repair cross complementing ). Среди них гены XPE и ERCC6-ERCC11 не требовались для прохождения основных реакций эксцизионной репарации, и их функция неизвестна.
Эксцизионная репарация нуклеотидов (ЭРН) E. coli
У E. coli эксцинуклеаза формируется, в результате димеризации двух молекул белка UvrА в присутствии АТР и связывания с одной молекулой UvrB. Гетеротример UvrA2B связывается с ДНК и с помощью 5′-3′-ДНК-геликазной активности перемещается вдоль ДНК в поисках повреждения. Такова предполагаемая картина узнавания эксцинуклеазой UvrAВС неспецифического повреждения в ДНК [ Hoeijmakers J., 1993 ]. Результатом узнавания повреждения этим комплексом явится внедрение субъединицы UvrB в ДНК, сопровождающееся конформационными изменениями ДНК в сайте внедрения ( изломы , локальная денатурация), диссоциация обеих субъединиц UvrА из комплекса и последующее связывание с ним молекул UvrD и UvrC. Гетеродимер UvrBC из состава нового комплекса делает два разрыва в поврежденной нити ДНК на расстоянии 8 н. с 5′-конца от повреждения и 5 н. с 3′-конца, катализируемых субъединицами UvrC и UvrB соответственно, что приводит к появлению 12-13-мерного олигонуклеотида,
вытесняемого из комплекса с помощью геликазы UvrD. Образующаяся при этом брешь заполняется ДНК-полимеразой I, синтезирующей ДНК по неповрежденной матрице, и сшивается ДНК-лигазой. Описанный процесс зависит от АТР. Действительно, АТР стимулирует димеризацию молекул UvrA, гидролиз АТР необходим для образования комплекса UvrA2B и проявления его геликазной активности, АТР необходим для формирования комплекса UvrB-ДНК и, наконец, для описанной выше бимодальной инцизии ДНК [ Friedberg E.C., Walker G.C., ea., 1991 ]. У E. coli транскрипционно-зависимую ветвь ЭРН (ТЭРН) осуществляет продукт гена mfd (mutation frequency decline). Этот ген был открыт за 35 лет до того, как его продукт признали фактором TRCF. Последний способствует диссоциации РНК-полимеразы от дефектной транскрибируемой нити ДНК, и связывается с субъединицей UvrA эксцинуклеазного комплекса. Иными словами, именно белок Mfd отыскивает повреждения на транскрибируемой нити и направляет ТЭРН на эту мишень.
Согласно недавним наблюдениям, два главных белка системы ДКНО — MutL и MutS — необходимы для ТЭРН [106 Melon I., Champl G.N., 1996106]. Причина такой взаимосвязи систем ДКНО и ТЭРН или заимствования белков MutL и MutS для выполнения сходных функций пока остается загадочной, хотя сам факт несомненен и нашел подтверждение также и для белков Msh2, Mlh1, Pms1 и Msh3 у S. cerevisiae [ Sweder K.S., ea, 1996 ].
Эксцизионная репарация нуклеотидов (ЭРН) у человека
Большому прогрессу в раскрытии механизма ЭРН у эукариот мы обязаны наследственному заболеванию человека — пигментной ксеродерме .
Изучение молекулярных основ этого заболевания выявили их сопряженность с дефектами системы ЭРН, результатом чего и явилось раскрытие генетического контроля ЭРН. Комплементационный анализ различных клеточных линий ХР определил 8 комплементационных групп (7 от XPA до XPG и одна «вариантная форма» XPV) [ Friedberg E.C., Walker G.C., ea., 1991 ]), а комплементационная
коррекция возможных мутантных линий клеток китайского хомячка, чувствительных к УФ-свету, способствовала выявлению дополнительных генов системы ЭРН. Последние получили название гены ERCC (excision repair cross-complimenting), и поэтому в названии генов и белков ЭРН используются обе аббревиатуры. Хотя детали процесса ЭРН у человека не ясны, так как не определена роль целого ряда генов, в первом приближении он выглядит так ( табл. 3 ) [ Lehmann A.R., 1995 , Lindahl T., ea, 1997 , Hoeijmakers J., 1993 ]. Белок ХРА в комплексе с онДНК-связующим белком RPA ( репликационным белком А ), транслоцируясь вдоль онДНК, опознает конформационнoе повреждениe. Взаимодействуя через другой участок белка ХРА с базальным фактором транскрипции TFIIH (две из субъединиц которого, белки ХРВ и ХРD , обладают геликазной активностью с противоположной ориентацией раскручивания днДНК), они образуют комплекс, который расплетает ДНК вокруг повреждения (в состав этого комплекса входит и белок ХРС
с неясными функциями). Через третий участок белка ХРА к комплексу примыкает гетеродимер ERCC1-XPF , который вносит однонитевой разрыв в ДНК с 5′-конца на расстоянии 16-25 н. от повреждения, тогда как белок XPG , входящий в комплекс белков эксцинуклеазы через взаимодействие с белком RPA, делает надрез с 3′-конца на расстоянии 2-9 н. (в разрезании принимает участие и белок ХРС , а белок ХРЕ активирует реакцию). В результате бимодальной инцизии участок ДНК размером около 29 н. высвобождается, а образующаяся брешь ресинтезируется с помощью ДНК-полимеразы епсилон или ДНК-полимеразы дельта , сопутствующего репликации фактора PCNА , репликационного фактора C-RFС и ДНК-лигазы I .
Представленная картина во многом основана на реконструировании процесса в открытой системе с участием 10 хорошо очищенных белков [ Wood R.D., 1994 ]. Как видно, эукариотическая система ЭРН лишь функционально подобна прокариотической. В ней задействовано значительно больше белков, число которых должно еще возрасти
за счет белков, осуществляющих разборку и сборку хроматина. Раскрытие природы ТЭРН у человека также связывают с пониманием молекулярного дефекта,приводящего к развитию двух мультисистемных генетических заболеваний — синдрома Кокейна (СS) и трихотиодистрофии ( ТТD ). При обоих заболеваниях соматические клетки больных оказались неспособны к ТЭРН. Классические случаи CS оказались связанными с повреждениями в двух генах, CSA и CSB, а редкие смешанные формы CS+XP (CS с дополнительными симптомами ХР), с повреждениями в генах ХРВ, ХРD и ХРG. При ТТD дефекты были обнаружены также в генах XPB и XPD и в новом пока не клонированном гене ТТDА, продукт которого является частью корового домена транскрипционного фактора TFIIH [ Lehmann A.R., 1995 , Lindahl T., ea, 1997 ].
Фактор TFIIH состоит из 6 субъединиц, две из которых — белки XPB и XPD. Связываясь с белками РНК-полимеразного комплекса, кор TFIIH принимает участие в инициировании транскрипции [ Buratowski S., 1994 ], a вкупе с белками репарационного
комплекса кор этого фактора участвует в ЭРН. Если допустить, что белки CSA и CSB способствуют превращению транскрипционного комплекса в репарационнный [ Lindahl T., ea, 1997 , Bregman D.B., 1996 ], то следствием повреждения этих белков может стать дефектность в ТЭРН. Таким образом, либо прямыми воздействиями на коровый домен фактора TFIIH (мутации в генах XPB, XPD и TTDA), либо косвенными (через гены CSA и CSB) можно модифицировать способность этого фактора к диссоциации из РНК- полимеразного комплекса для участия в ТЭРН. Что касается белка XPG, то он, подобно своему гомологу из S. cereivisiae — белку Rad2 [ Bregman D.B., 1996 ], способен взаимодействовать c TFIIH, что допускает возможность его воздействия на TFIIH для участия в ТЭРН. Таковы гипотетические объяснения взаимосвязи молекулярных дефектов при CS и ТTD с нарушениями ТЭРН, которые могут быть полезными для раскрытия механизма ТЭРН у эукариот. Связь же между молекулярными дефектами и клиническими проявлениями
рассматриваемых наследственных заболеваний пока не имеет даже упрощенных толкований [ Kolodner R., 1995 ].
Так случилось, что изучение молекулярных механизмов ЭРН и ТЭРН на клетках человека развивались параллельно, или даже опережая исследования на популярной эукариотической модели дрожжей S. cerevisiae. В табл. 3 представлены известные функциональные аналоги системы ЭРН у дрожжей. Несомненно, однако, что при исследовании детального механизма ЭРН именно эта модельная система будет играть ведущую роль. Например, немаловажной особенностью системы ЭРН у E. coli является ее связь с SOS-функциями клетки. Действительно, центральные гены системы ЭРН, uvrA и uvrB, находятся под контролем SOS-регулона [ Friedberg E.C., Walker G.C., ea., 1991 ]. Системы, подобной SOS, у эукариот пока не обнаружено. Однако целый ряд генов системы ЭРН у дрожжей, таких, как RAD2, RAD7, RAD23, CDC8 и CDC9, индуцируется при повреждении ДНК, а последние два гена дополнительно регулируются клеточным циклом [ Friedberg
E.C., Walker G.C., ea., 1991 ].
Природу этой индукции еще предстоит выяснить.
Текст работы размещён без изображений и формул.
Полная версия работы доступна во вкладке «Файлы работы» в формате PDF
Актуальность: Данная тема является актуальной по причине того, что репарация является свойством живой клетки бороться с различными повреждениями ДНК, чтобы сохранить целостность ДНК. Данная тема затрагивает одну из важных задач современной медицины.
Цель: понять, как нарушается целостность генетического материала клетки и какие существуют восстановительные механизмы ДНК и как они работают.
Задачи:
1. Изучить, что такое репарация.
2. Разобрать, как происходит повреждение ДНК и по каким причинам
3. Выяснить, как работают репарационные механизмы ДНК
Содержание………………………………………………………………….стр.
1. Введение………………………………………………………………….…3
2. Нарушения в ДНК…………………………..…………………………….…4
3. «Световое восстановление»………………………………………….……..5
4. «Темновое восстановление»……………………………………………….6
5. Система репарации………………………………………………….……..7
6. Классификация репарации………………………………………………..8
7. Виды репарации……………………………………………………………9
8. Интересные факты о репарации………………………………….………11
9. Заключение………………………………………………………………..12
10. Список литературы………………………………………………………13
Введение
Репарация (от лат. reparatio — восстановление) — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённых при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических реагентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней (напр., пигментная ксеродерма) связан с нарушениями систем репарации. В окружающем мире существует множество факторов, способных вызвать необратимые изменения в живом организме. Чтобы сохранить свою целостность, избежать патологических и несовместимых с жизнью мутаций, должна существовать система самостоятельного восстановления.
Нарушения в ДНК
Молекула дезоксирибонуклеиновой кислоты может быть разорвана как в ходе биосинтеза, так и под влиянием вредных веществ.
Источники повреждения ДНК:
— Ультрафиолетовое излучение
— Радиация
— Химические вещества
— Ошибки репликации ДНК
— Апуринизация — отщепление азотистых оснований от сахарофосфатного остова
— Дезаминирование — отщепление аминогруппы от азотистого основания
К негативным факторам, в частности, относят температуру или физические силы различного происхождения. Если разрушение произошло, клетка запускает процесс репарации. Так начинается восстановление исходной структуры молекулы ДНК. За репарацию отвечают особые ферментные комплексы, присутствующие внутри клеток. С невозможностью отдельных клеток осуществлять восстановление связаны некоторые заболевания.
«Световое восстановление»
Впоследствии исследования Кельнера получили свое логическое продолжение в работах американских биологов Сетлоу, Руперта и некоторых других. Благодаря труду этой группы ученых было достоверно установлено, что фотореактивация является процессом, который запускается благодаря особому веществу – ферменту, катализирующему расщепление димеров тимина. Именно они, как выяснилось, образовывались в ходе экспериментов под воздействием ультрафиолета. При этом яркий видимый свет запускал действие фермента, который способствовал расщеплению димеров и восстановлению первоначального состояния поврежденных тканей. В данном случае речь идет о световой разновидности восстановления ДНК. Определим это более четко. Можно сказать, что световая репарация – это восстановление под воздействием света первоначальной структуры ДНК после повреждений. Однако данный процесс не является единственным, способствующим устранению повреждений.
«Темновое восстановление»
Спустя некоторое время после открытия световой была обнаружена темновая репарация. Это явление происходит без какого-либо воздействия световых лучей видимого спектра. Данная способность к восстановлению обнаружилась во время исследования чувствительности некоторых бактерий к ультрафиолетовым лучам и ионизирующему излучению. Темновая репарация ДНК – это способность клеток убирать любые патогенные изменения дезоксирибонуклеиновой кислоты. Но следует сказать, что это уже не фотохимический процесс, в отличие от светового восстановления.
Механизм «темнового» устранения повреждений Наблюдения за бактериями показали, что спустя некоторое время после того, как одноклеточный организм получил порцию ультрафиолета, вследствие чего некоторые участки ДНК оказались поврежденными, клетка регулирует свои внутренние процессы определенным образом. В результате измененный кусочек ДНК просто отрезается от общей цепочки. Получившиеся же промежутки заново заполняются необходимым материалом из аминокислот. Иными словами, осуществляется ресинтез участков ДНК. Открытие учеными такого явления, как темновая репарация тканей, – это еще один шаг в изучении удивительных защитных способностей организма животного и человека.
Система репарации
Эксперименты, позволившие выявить механизмы восстановления и само существование этой способности, проводились с помощью одноклеточных организмов. Но процессы репарации присущи живым клеткам животных и человека. Некоторые люди страдают пигментной ксеродермой. Это заболевание вызвано отсутствием способности клеток ресинтезировать поврежденную ДНК. Ксеродерма передается по наследству. Из чего же состоит репарационная система? Четыре фермента, на которых держится процесс репарации – это ДНК-хеликаза, -экзонуклеаза, -полимераза и -лигаза. Первый из этих соединений способен распознавать повреждения в цепи молекулы дезоксирибонуклеиновой кислоты. Он не только распознает, но и обрезает цепь в нужном месте, чтобы удалить измененный отрезок молекулы. Само устранение осуществляется с помощью ДНК-экзонуклеазы. Далее происходит синтез нового участка молекулы дезоксирибонуклеиновой кислоты из аминокислот с целью полностью заменить поврежденный отрезок. Ну и финальный аккорд этой сложнейшей биологической процедуры совершается с помощью фермента ДНК-лигазы. Он отвечает за прикрепление синтезированного участка к поврежденной молекуле. После того как все четыре фермента сделали свою работу, молекула ДНК полностью обновлена и все повреждения остаются в прошлом. Вот так слаженно работают механизмы внутри живой клетки.
Классификация репарации
На данный момент ученые выделяют следующие разновидности систем репарации. Они активируются в зависимости от разных факторов. К ним относятся: реактивация, рекомбинационное восстановление, репарация гетеродуплексов, эксцизионная репарация, воссоединение негомологичных концов молекул ДНК. Все одноклеточные организмы обладают как минимум тремя ферментными системами. Каждая из них обладает способностью осуществлять процесс восстановления. К этим системам относят: прямую, эксцизионную и пострепликативную. Этими тремя видами восстановления ДНК обладают прокариоты. Что касается эукариот, то в их распоряжении находятся дополнительные механизмы, которые называются Miss-mathe и Sos-репарация. Биология подробно изучила все эти виды самовосстановления генетического материала клеток.
Виды репарации
1. Прямая репарация — наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов. Так действует, например, O6-метилгуанин-ДНК-метилтрансфераза, которая снимает метильную группу с азотистого основания на один из собственных остатков цистеина.
2. Эксцизионная репарация (англ. excision — вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы. Эксцизионная репарация (excision repair): процесс с участием ферментативной системы, которая удаляет короткую однонитевую последовательность двунитевой ДНК , содержащей ошибочно спаренные или поврежденные основания , и замещает их путем синтеза последовательности, комплементарной оставшейся нити. Эксцизионная репарация является наиболее распространенным способом репарации модифицированных оснований ДНК. Этот тип репарации базируется на распознавании модифицированного основания различными гликозилазами, расщепляющими N-гликозидную связь этого основания с сахарофосфатным остовом молекулы ДНК. При этом существуют гликозилазы, специфически распознающие присутствие в ДНК определенных модифицированных оснований (оксиметилурацила, гипоксантина, 5-метилурацила, 3-метиладенина, 7-метилгуанина и т.д.). Для многих гликозилаз к настоящему времени описан полиморфизм, связанный с заменой одного из нуклеотидов в кодирующей последовательности гена. Для ряда изоформ этих ферментов была установлена ассоциация с повышенным риском возникновения онкологических заболеваний.
3. Пострепликативная репарация Tип репарации, имеющей место в тех случаях, когда процесс эксцизионной репарации недостаточен для полного исправления повреждения: после репликации с образованием ДНК, содержащей поврежденные участки, образуются одноцепочечные бреши, заполняемые в процессе гомологичной рекомбинации при помощи белка. Пострепликативная репарация была открыта в клетках E.Coli, не способных выщеплять тиминовые димеры. Это единственный тип репарации, не имеющий этапа узнавания повреждения.
Интересные факты о репарации
1. Полагают, что от 80 % до 90 % всех раковых заболеваний связаны с отсутствием репарации ДНК[4].
2. Повреждение ДНК под воздействием факторов окружающей среды, а также нормальных метаболических процессов, происходящих в клетке, происходит с частотой от нескольких сотен до 1000 случаев в каждой клетке, каждый час.
3. По сути ошибки в репарации происходят так же часто как и в репликации, а при некоторых условиях даже чаще.
4. В половых клетках сложная репарация, связанная с гомологичной рекомбинацией не происходит из-за гаплоидности генома этих клеток.
Заключение
Репарация – это обязательное условие нормального функционирования организма. Подвергаясь ежедневно и ежечасно угрозам повреждений и мутаций ДНК, многоклеточная структура приспосабливается и выживает. Это происходит в том числе и за счет налаженной системы репарации. Отсутствие нормальной восстановительной способности вызывает болезни, мутации и другие отклонения. К ним относятся различные патологии развития, онкология и даже само старение. Наследственные болезни вследствие нарушений репарации могут приводить к тяжелым злокачественным опухолям и другим аномалиям организма. Сейчас определены некоторые заболевания, вызываемые именно сбоями систем репарации ДНК. Это такие, например, патологии, как синдром Кокейна, ксеродерма, неполипозный рак толстой кишки, трихотиодистрофия и некоторые раковые опухоли.
Список литературы
1. Кирпичев А.С., Севастьянова Г.А. Молекулярная биология М. AKADEMA. 2005г.
2. С.Коничев, Г.А.Севастьянова Молекулярная биология. — Москва: Академия, 2003г.
3. С.Г. Инге-Вечтомов. Генетика с основами селекции. — Москва: Высшая школа, 1989г.
4. Кирилл Стасевич Как клетка чинит свою ДНК. Наука и жизнь. — 2015г.
5. Б. Льюин. Гены 1987г.
Репарация
(от лат. reparatio — восстановление) — особая
функция клеток, заключающаяся в
способности исправлять химические
повреждения и разрывы в молекулах ДНК,
повреждённой при нормальном биосинтезе
ДНК в клетке или в результате воздействия
физических или химических агентов.
Осуществляется специальными ферментными
системами клетки. Ряд наследственных
болезней (напр., пигментная ксеродерма)
связан с нарушениями систем
репарации.
(Процесс,
позволяющий живым организмам
восстанавливать повреждения, возникающие
в ДНК, называют репарацией – кратко).
Все
репарационные механизмы основаны на
том, что ДНК — двухцепочечная молекула,
т.е. в клетке есть 2 копии генетической
информации. Если нуклеотидная
последовательность одной из двух цепей
оказывается повреждённой (изменённой),
информацию можно восстановить, так как
вторая (комплементарная) цепь
сохранена.
Процесс
репарации происходит в несколько этапов.
На первом этапе выявляется нарушение
комплементарности цепей ДНК. В ходе
второго этапа некомплементарный
нуклеотид или только основание
устраняется, на третьем и четвёртом
этапах идёт восстановление целостности
цепи по принципу комплементарности.
Однако в зависимости от типа повреждения
количество этапов и ферментов, участвующих
в его устранении, может быть разным.
Очень
редко происходят повреждения, затрагивающие
обе цепи ДНК, т.е. нарушения структуры
нуклеотидов комплементарной пары. Такие
повреждения в половых клетках не
репарируются, так как для осуществления
сложной репарации с участием гомологичной
рекомбинации требуется наличие
диплоидного набора хромосом.
Виды:
1)Прямая
репарация
2)Эксцизионная
репарация
3)Пострепликативная
репарация
Прямая
репарация
— наиболее простой путь устранения
повреждений в ДНК, в котором обычно
задействованы специфические ферменты,
способные быстро (как правило, в одну
стадию) устранять соответствующее
повреждение, восстанавливая исходную
структуру нуклеотидов. Так действует,
например, O6-метилгуанин-ДНК-метилтрансфераза,
которая снимает метильную группу с
азотистого основания на один из
собственных остатков цистеина.
Эксцизионная
репарация (англ. excision — вырезание)
включает удаление повреждённых азотистых
оснований из ДНК и последующее
восстановление нормальной структуры
молекулы. Эксцизионная репарация
(excision repair): процесс с участием ферментативной
системы, которая удаляет короткую
однонитевую последовательность
двунитевой ДНК , содержащей ошибочно
спаренные или поврежденные основания
, и замещает их путем синтеза
последовательности, комплементарной
оставшейся нити.
Эксцизионная
репарация является наиболее распространенным
способом репарации модифицированных
оснований ДНК. Этот тип репарации
базируется на распознавании
модифицированного основания различными
гликозилазами, расщепляющими N-гликозидную
связь этого основания с сахарофосфатным
остовом молекулы ДНК. При этом существуют
гликозилазы, специфически распознающие
присутствие в ДНК определенных
модифицированных оснований
(оксиметилурацила, гипоксантина,
5-метилурацила, 3-метиладенина,
7-метилгуанина и т.д.). Для многих гликозилаз
к настоящему времени описан полиморфизм,
связанный с заменой одного из нуклеотидов
в кодирующей последовательности гена.
Для ряда изоформ этих ферментов была
установлена ассоциация с повышенным
риском возникновения онкологических
заболеваний [ Chen, 2003 ].
Пострепликативная
репарация
Tип
репарации, имеющей место в тех случаях,
когда процесс эксцизионной репарации
недостаточен для полного исправления
повреждения: после репликации с
образованием ДНК, содержащей поврежденные
участки, образуются одноцепочечные
бреши, заполняемые в процессе гомологичной
рекомбинации при помощи белка
RecA.[1]
Пострепликативная
репарация была открыта в клетках E.Coli,
не способных выщеплять тиминовые димеры.
Это единственный тип репарации, не
имеющий этапа узнавания повреждения.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Содержание
- Что такое механизм репарации ДНК?
- Типы механизма репарации ДНК
- 1. Система устранения несоответствия
- 2. Иссечение основания
- 3. Восстановление эксцизии нуклеотидов
- 4. Прямой ремонт
Что такое механизм репарации ДНК?
Ремонт ДНК — это совокупность процессов, с помощью которых клетка идентифицирует и исправляет повреждение молекул ДНК, кодирующих ее геном.
определение ремонта ДНК
- В каждой клетке обычно присутствует один или два набора геномной ДНК. Информация ДНК может быть использована для быстрой замены сломанных белков и молекул РНК, но молекулы ДНК не могут быть восстановлены.
- Сложная система механизмов репарации ДНК поддерживает клеточную потребность в поддержании целостности информации в ДНК.
- Многочисленные механизмы, как спонтанные, так и вызванные факторами окружающей среды, могут разрушать ДНК.
- Информационное содержание ДНК иногда может быть повреждено самой репликацией, если ошибки приводят к несовпадению пар оснований (например, G в паре с T).
- Повреждение ДНК имеет разнообразную и сложную химию. Многочисленные ферментные системы, которые катализируют некоторые из самых интересных химических изменений в метаболизме ДНК, являются частью клеточного ответа на это повреждение.
- Сначала мы рассмотрим результаты изменений последовательности ДНК, прежде чем думать о конкретных механизмах восстановления.
Типы механизма репарации ДНК
Существует четыре типа системы восстановления ДНК, такие как;
- Исправление несоответствия
- Иссечение основания
- Нуклеотид-эксцизионная репарация
- Прямой ремонт
1. Система устранения несоответствия
- Исправление нескольких несоответствий, оставшихся после репликации в E. coli, повышает общую точность репликации в 10^2–10^3 раз.
- Несоответствия почти всегда исправляются, чтобы отразить информацию в старой (шаблонной) цепочке; следовательно, механизм репарации должен различать матрицу и вновь синтезированную цепь.
- Клетка достигает этого, присоединяя метильные группы к матричной ДНК, чтобы отличить ее от вновь продуцируемых цепей.
- Система репарации несоответствия E. coli состоит как минимум из 12 белковых компонентов, которые участвуют либо в разделении цепей, либо в самом процессе репарации.
- Механизм, лежащий в основе дискриминации нитей у большинства бактерий и эукариот, не выяснен, однако он хорошо изучен у E. coli и некоторых близкородственных бактерий.
- Dam метилаза, которая, как вы помните, метилирует ДНК в Af-положении всех аденинов внутри (5′)GATC последовательностей, ответственна за различение цепей у этих бактерий.
- Существует короткий интервал (несколько секунд или минут) после прохождения репликационной вилки, в течение которого матричная цепь метилируется, а свежесинтезированная цепь — нет.
- Временный неметилированный статус последовательностей GATC в свежесинтезированной цепи отличает вновь синтезированную цепь от матричной цепи.
- Несоответствия репликации в области гемиметилированной последовательности GATC впоследствии репарируются с использованием информации из метилированной исходной цепи (матрицы).
- Если обе цепи метилированы в последовательности GATC, исправляются лишь некоторые несовпадения; если ни одна из цепей не метилирована, несоответствия исправляются, но ни одна из цепей не является предпочтительной.
- Из полуметилированной последовательности GATC метил-направленный механизм репарации несоответствий клетки эффективно устраняет несоответствия до 1,000 пар оснований.
Фермент участвует в процессе восстановления ДНК — восстановление несоответствия
| Ферменты/белки | Тип повреждения |
| Дамметилаза | |
| ДНК-лигаза | |
| Нуклеаза RecJ | |
| Экзонуклеаза X | |
| Экзонуклеаза VII | Несоответствия |
| Экзонуклеаза I | |
| ДНК-полимераза III | |
| SSB | |
| ДНК-хеликаза II | |
| Белки MutH, MutL, MutS |
Как процесс исправления несоответствия управляется относительно удаленными последовательностями GATC?
- Белок MutL объединяется с белком MutS с образованием комплекса, который связывает все несовпадающие пары оснований (кроме CC).
- Белок MutH связывается с последовательностями MutL и GATC, с которыми сталкивается комплекс MutL-MutS. ДНК с обеих сторон несоответствия проходит через комплекс MutL-MutS, образуя петлю ДНК.
- MutH обладает сайт-специфической эндонуклеазной активностью, которая неактивна до тех пор, пока не встретится полуметилированная последовательность GATC. В этом месте MutH катализирует расщепление неметилированной цепи 5′ до G в GATC, помечая ее для репарации.
- Следующие шаги пути зависят от расположения несоответствия относительно сайта расщепления.
- Когда несоответствие находится на 5′-стороне сайта расщепления, неметилированная цепь распутывается и разрушается от сайта расщепления через несоответствие, и этот участок заменяется новой ДНК.
- Эта процедура требует согласованных усилий ДНК-хеликазы II, SSB, экзонуклеазы I или экзонуклеазы X (обе из которых расщепляют нити ДНК в направлении 3′-5′), ДНК-полимеразы III и ДНК-лигазы.
- Экзонуклеаза VII разрушает одноцепочечную ДНК в направлении 5′-3′ или 3′-5′, тогда как нуклеаза RecJ разрушает одноцепочечную ДНК в направлении 5-3′.

Особенности системы восстановления несоответствия
- E. coli приходится тратить много энергии на исправление несоответствий, что является очень дорогим процессом.
- Разница может составлять не менее 1,000 пар оснований от последовательности GATC.
- Чтобы исправить одно несовпадающее основание, требуется много активированных предшественников дезоксинуклеотидов, чтобы расщепить и заменить сегмент нити этой длины.
- Это еще раз показывает, насколько важна целостность генома для клетки.
- Всего несколько белков эукариотические клетки которые структурно и функционально сходны с бактериальными белками MutS и MutL, но не с белком MutH.
- Изменения в генах человека, которые кодируют эти типы белков, вызывают некоторые из наиболее распространенных синдромов, повышающих вероятность развития рака у людей. Это еще раз показывает, насколько важны для организма системы репарации ДНК.
- Большинство эукариот, от дрожжей до человека, имеют MSH2, MSH3 и MSH6 в качестве основных гомологов MutS.
- Гетеродимеры MSH2 и MSH6 обычно связываются с петлями только с одной несовпадающей парой оснований и хуже связываются с петлями с двумя или более несовпадающими парами оснований.
- Во многих организмах более длинные несовпадения (2–6 п.н.) могут быть связаны гетеродимером MSH2 и MSH3, или оба типа гетеродимеров могут связываться с несовпадениями одновременно.
- Гомологи MutL, которые в основном состоят из MLHI (гомолог 1 MutL) и PMS1 (постмейотическая сегрегация), связываются с комплексами MSH и сохраняют их стабильность.
- В эукариотической репарации несоответствия еще много деталей, которые необходимо проработать. В частности, мы не знаем, как распознаются новообразованные нити ДНК, хотя исследования показали, что это не имеет ничего общего с последовательностью GATC.

2. Иссечение основания
- ДНК-гликозилазы — это тип фермента, который содержится в каждой клетке. Эти ферменты ищут обычные повреждения ДНК, такие как продукты дезаминирования цитозина и аденина, и удаляют поврежденное основание, разрывая N-гликозильную связь.
- Это расщепление создает в ДНК апуриновый или апиримидиновый сайт, также называемый АР-сайтом или абазовым сайтом.
- В большинстве случаев каждая ДНК-гликозилаза воздействует только на один тип поражения.
- Урацил-ДНК-гликозилазы, присутствующие в большинстве клеток, удаляют урацил из ДНК, образующейся при расщеплении цитозина.
- Многие мутации G:C в A=T происходят в клетках, которые потеряли этот фермент. Этот фермент не удаляет остатки урацила или тимина из РНК или ДНК.
- Способность различать тимин и урацил, возникающая в результате дезаминирования цитозина и необходимая для селективной репарации урацила, может быть одной из причин, по которой ДНК содержит тимин вместо урацила.
- У большинства бактерий есть только один вид урацил-ДНК-гликозилазы, но у людей есть как минимум четыре различных типа, что показывает, насколько важно избавиться от урацила из ДНК.
- Наиболее распространенная урацилгликозилаза у людей называется UNG. Он обнаружен в реплисоме человека, где он избавляется от случайного остатка U, который заменяет T во время репликации.
- Дезаминирование остатков C происходит в одноцепочечной ДНК в 100 раз быстрее, чем в двухцепочечной ДНК, и у людей есть фермент под названием hSMUGl, который избавляется от любых остатков U, возникающих во время репликации или транскрипции в одноцепочечной ДНК.
- Еще две ДНК-гликозилазы человека, TDG и MBD4, удаляют остатки U или T, которые спарены с G. Это происходит при дезаминировании цитозина или 5-метилцитозина.
- Другие ДНК-гликозилазы могут находить и избавляться от различных видов поврежденных оснований, таких как формамидопиримидин и 8-гидроксигуанин, образующиеся в результате окисления пуринов, гипоксантин, образующийся при дезаминировании аденина, и алкилированные основания, такие как 3-метиладенин и 7-метилгуанин.
- Также было обнаружено, что у некоторых типов организмов есть гликозилирующие ферменты, которые могут распознавать другие типы повреждений, такие как пиримидиновые димеры.
- Помните, что AP-сайты также создаются, когда N-гликозильные связи в ДНК медленно разрушаются сами по себе.
- После того, как ДНК-гликозилаза создает AP-сайт, его необходимо зафиксировать другим типом фермента.
- Вы не можете просто добавить новое основание и создать новую N-гликозильную связь, чтобы решить проблему. Вместо этого оставшийся дезоксирибозо-5′-фосфат удаляется, а на его место помещается новый нуклеотид.
- Этот процесс начинается с одного из ферментов, называемых АР-эндонуклеазами, который разрезает цепь ДНК в АР-сайте.
- В зависимости от типа АР-эндонуклеазы разрез может быть сделан на 5 или 3 футах от АР-сайта.
- Затем AP-сайт вырезается из фрагмента ДНК, который заменяется ДНК-полимеразой I, а оставшийся разрыв запечатывается ДНК-лигазой.
- У эукариот замена нуклеотидов осуществляется полимеразами, предназначенными для этой задачи.

Фермент участвует в процессе восстановления ДНК – Base-Excision Repair
| Ферменты и белки | Тип поврежденияe |
| ДНК-гликозилазы | |
| AP эндонуклеазы | Аномальные основания (урацил, гипоксантин, ксантин); алкилированные основания; в некоторых других организмах пиримидиновые димеры |
| ДНК-полимераза I | |
| ДНК-лигаза |
3. Восстановление эксцизии нуклеотидов
- Система удаления нуклеотидов — это путь репарации, важный для выживания всех свободноживущих организмов. Он исправляет повреждения ДНК, которые вызывают большие изменения в спиральной структуре ДНК.
- При репарации с иссечением нуклеотидов мультисубъединичный фермент, называемый эксинуклеазой, расщепляет две фосфодиэфирные связи, по одной с каждой стороны повреждения, вызванного поражением.
- У E. coli и других бактерий ферментная система разрывает пятую фосфодиэфирную связь на 3′-стороне и восьмую фосфодиэфирную связь на 5′-стороне. Это составляет фрагмент из 12–13 нуклеотидов (в зависимости от того, затрагивает ли повреждение одно или два основания).
- У человека и других эукариот ферментная система разрывает шестую фосфодиэфирную связь на 3′-стороне и двадцать вторую фосфодиэфирную связь на 5′-стороне. Это составляет фрагмент от 27 до 29 нуклеотидов.
- После двойного разреза удаленные олигонуклеотиды освобождаются от дуплекса, и этот пробел заполняет ДНК-полимераза I E. coli или ДНК-полимераза человека. Разрыв закрывается ДНК-лигазой.
- В E. coli наиболее важным ферментным комплексом является эксинуклеаза ABC. Он состоит из трех частей: UvrA (M.104,000 78,000), UwB (M.68,000 XNUMX) и UwC. (М.XNUMX).
- Название «эксинуклеаза» происходит от того факта, что этот ферментный комплекс является единственным, который может катализировать два специфических эндонуклеолитических расщепления. Это отличает ее от других эндонуклеаз.
- Комплекс белков UwA и UwB, называемый A2B, смотрит на ДНК и связывается с поврежденным участком.
- Затем димер UwA распадается, оставляя плотный комплекс UwB и ДНК.
- Затем белок UwC связывается с UwB, а UwB разрезает пятую фосфодиэфирную связь на 3′-стороне поражения. После этого UvrC разрезает восьмую фосфодиэфирную связь на 5′-стороне.
- Затем геликаза UvrD избавляется от оставшегося фрагмента длиной от 2 до 13 нуклеотидов.
- ДНК-полимераза I и ДНК-лигаза затем заполняют оставшийся короткий пробел.
- Этот путь является основным способом фиксации многих типов повреждений ДНК, таких как димеры циклобутан-пиримидина, 6-4-фотопродукты и несколько типов аддуктов оснований, таких как бензо[о]пирен-гуанин, который образуется в ДНК при подвергается воздействию сигаретного дыма.
- Эксинуклеаза ABC обладает уникальной нуклеолитической активностью, заключающейся в том, что она делает два разреза в ДНК.
- Принцип работы эукариотических экзонуклеаз аналогичен действию бактериального фермента, но для двойного разреза требуется 16 полипептидов, не имеющих ничего общего с субъединицами эксинуклеазы E. coli.
- У эукариот транскрипция тесно связана как с репарацией с удалением нуклеотидов, так и с репарацией с удалением оснований. Люди с проблемами в генах эксцизионной репарации нуклеотидов с большей вероятностью получат ряд серьезных заболеваний.

Фермент участвует в процессе репарации ДНК – репарация нуклеотидов-эксцизия
| Ферменты/белки | Тип повреждения |
| ABC эксинуклеаза | |
| ДНК-полимераза I | Повреждения ДНК, вызывающие большие структурные изменения (например, пиримидиновые димеры) |
| ДНК-лигаза |
4. Прямой ремонт

- Существуют различные виды повреждений, которые можно исправить, не удаляя основание или нуклеотид. ДНК-фотолиазы ускоряют прямую фотореактивацию димеров циклобутан-пиримидина, что является наиболее известным примером.
- Ультрафиолетовый свет вызывает реакцию, в результате которой образуются пиримидиновые димеры, а фотолиазы используют энергию поглощенного света для устранения повреждений.
- Большинство фотолиаз имеют два кофактора, которые действуют как хромофоры или вещества, поглощающие свет. ФАДГ- всегда является одним из хромофоров.

- Другой цвет кишечной палочки и дрожжей — это фолиевая кислота. В ходе реакции образуются свободные радикалы.
- ДНК-фотолиазы не обнаружены в клетках млекопитающих, рожающих в плаценте (включая человека). В репарации нуклеотидов с повреждением алкилирования вы можете увидеть больше примеров этого.
- При наличии алкилирующих агентов образуется модифицированный нуклеотид O6-метилгуанин. Это распространенное и очень мутагенное поражение. Он имеет тенденцию образовывать пары с тимином вместо цитозина во время репликации, что вызывает переход от G:C к A:T.

- O6-метилгуанин-ДНК-метилтрансфераза представляет собой белок, который помогает перемещать метильную группу O6-метилгуанина к одному из его собственных остатков Cys. Так осуществляется прямая репарация 06-метилгуанина.
- Эта метилтрансфераза на самом деле не является ферментом, потому что одиночный перенос метила постоянно метилирует белок, делая его неактивным в этом пути.

- Еще один яркий пример того, насколько важно поддерживать клеточную ДНК в хорошем состоянии, заключается в том, что для фиксации одного сломанного основания требуется целая белковая молекула.
- 1-метиладенин и 3-метилцитозин фиксируются совершенно другим, но все же прямым способом.
- Когда ДНК является одноцепочечной, аминогруппы остатков А и С иногда метилированы. Это имеет прямое влияние на то, насколько хорошо пары оснований.
- В E. coli белок AlkB, который является членом надсемейства о-кетоглутарат-Fe2+-зависимых диоксигеназ, отвечает за окислительное деметилирование этих алкилированных нуклеотидов.

Фермент участвует в процессе восстановления ДНК — прямое восстановление
| Фермент | Тип повреждения |
| фотолиазы ДНК | |
| О6-метилгуанин-ДНК-метилтрансфераза | Пиримидиновые димеры 06-метилгуанин |
| Белок AlkB | 1-метилгуанин, 3-метилцитозин |
Отправьте свой вопрос
Пожалуйста, задайте свой вопрос в соответствующей категории.
Похожие темы
рис.1 Модифицированные азотистые основания ДНК, удаляемые ДНК–гликозилазами: а – урацил; б – гипоксантин; в – 5–гидроксицитозин; г – 2,5-диамино-4-формамидопиримидин; д – 7,8-дигидро-8-оксогуанин; е – мочевина; ж – тимингликоль; з – 5-формилурацил; и – 5-гидроксиметилурацил; к – 3-метиладенин; л – 7-метилгуанин; м – 2-метилцитозин [Партушев, 2000].
АP-эндонуклеаза создает ник (одноцепочечный разрав) с 3’-ОН и 5’-dRP концами. Для большинства млекопитающих характерен
тип BER репарации с включением одного нуклеотида. В этом случае ДНК-полимераза β вставляет 1 нуклеотид в 3’-конец праймера и затем удаляет 5’-dRP фрагмент с помощью своей dRP-лиазной активности. Получившийся ник сшивается ДНК-лигазой. Альтернативным является путь так называемой длинно-заплаточной BER репарации. Он реализуется в тех случаях, когда 5’-dRP фрагмент модифицирован и не может быть удален с помощью ДНК–полимеразы β. В этом случае Pol β проводит синтез ДНК с вытеснением запирающей цепи, 2 — 13 нуклеотидов. Образующийся в результате свисающий 5’-конец ДНК выщепляется флэпэндонуклеазой FEN1. Pol бета, Pol дельтаи Pol епсилон могут вести ситнез ДНК во время длинно-заплаточной репарации. Идентичность полимераз, вовлеченных в этот процесс in vitro, еще не ясна, но показано, что Pol β всегда инициирует синтез ДНК.
Зависимость того, какой из путей BER реализуется, определяется бифункциональными ДНК-гликозилазами, которые имеют дополнительную
АP-лиазную активность. При комбинации ДНК-гликозилазной, АP-лиазной и АP-эндонуклеазной активностей внутри одного фермента в поврежденной цепи ДНК образуется однонуклеотидная брешь с 3’-ОН и 5’-P концами, которую может застроить Pol β.
Дополнительная сложность в понимании механизмов переключения внутри BER состоит в том, что две новые полимеразы Pol i и Pol лямбда также имеют dRP-лиазнуюактивность. Таким образом, можно предположить, что они, как и Pol бета, являются участниками тех процессов репарации, где необходимо выщепление dRP фрагмента. Pol лямбда является близким гомологом Pol бета со сходными ферментативными свойствами. Также как и Pol бета, она лишена 3’—>5’экзонуклеазной активности и имеет низкую процессивность синтеза на частичном ДНК-дуплексе, содержащем свисающий 5’-участок матрицы, процессивный синтез возможен в брешах с 5’-P концами. Таким образом, Pol лямбда является подходящим кандидатом для BER синтеза. Более того,
Pol лямбда заменяет Pol бета в реконструированных BER системах, репарирующих урацил-содержащие ДНК, iv vitro. Однако, клетки мышей Pol лямбда -/- не чувствительны к обработке перекисью или метилметансульфонатом, агентам, которые, помимо других типов повреждений, продуцируют АР-сайты и окисленные формы оснований. Отсюда можно заключить, что Pol лямбда не является необходимой для BER в клетках, где есть, Pol дельта и Pol епсилон. Интересно, что Pol лямбда может эффективно процессировать ДНК при очень низкой концентрации dNTP (около 1 мкМ), что может означать ее участие в любых клеточных процессах в фазе G0 при низкой концентрации dNTP. В поддержку этой гипотезы выступает тот факт, что экспрессия Pol лямбда зависит от клеточного цикла, и наибольшее количества белка экспрессируется при переходе из S- в М-фазу и в спокойных клетках.
Pol i принадлежит к Y семейству полимераз. Основная функция полимераз этого семейства – утилизация ДНК-повреждений, блокирующих работу репликативных ДНК-полимераз. Однако,
исходя из некоторых свойств Pol i, можно предположить, что этот фермент является участником альтернативного процесса BER репарации. Pol i обладает низкой поцессивностью, лишена 3’>5’экзонуклеазной активности и способна застраивать брешь в 1 — 5 нуклеотидов iv vitro. Pol i может замещать Pol бета в реконструированных системах BER, репарирующих урацил-содержащие ДНК, iv vitro, благодаря наличию dRP-лиазной и ДНК-полимеразной активностей. Кинетические исследования реакции нуклеотидного встраивания овыявили дополнительные данные, свидетельствующие в пользу того, что Pol i может принимать участие в репарации. Pol i вставляет dTMP напротив А в ДНК-матрицу с большей эффективностью, чем любой другой дезоксинуклеозидмонофосфат, при этом точность ДНК-синтеза сопоставима с параметрами для Pol бета. Основываясь на этих данных, можно предположить, что Pol i – участник BER репарации оснований уридина, который образуется после встраивания dUMP напротив А во время ошибочной репликации. Pol i также
может вставлять dGМP напротив Т со скоростями, близкими к скорости включения корректного нуклеотида. Более того, на матрице, содержащей 2 или более последовательных Т, вторым встраиваемым нуклеотидом оказался dGMP. Подобные результаты служат основой для гипотезы, что Pol i может быть участником альтернативной BER репарации в тех слсучаях, когда dG был ошибочно удален гликозилазой из G-T или G-U некомплементарной пары, образовавшейся в результате дезаминации 5-метилцитозина или цитозина. Возможная роль Pol i в трансляции синтеза ДНК и процессе соматических гипермутаций рассмотрена ниже.
Возможность участия пяти ядерных полимераз в BER, три из которых обладают dRP-лиазной активностью, была мало изучена в клетках и на модельных животных, лишенных более чем одной полимеразы. Поэтому данные о субстратной специфичности и взаимодействии полимеразо-акцепторных белков, необходимых в BER, недостаточны. Помимо BER репарации, Pol бетта принимает участие в репарации одноцепочечных разрывов; функционирование
этого процесса нарушено у пациентов с наследственной спинномозговой атаксией. Одноцепочечные разрывы генерируются эндогенными или экзогенными агентами. Такие разрывы часто содержат однонуклеотидные бреши с 3’ и/или 5’ модифицированными концами, то есть очень похожи на субстраты BER. Так вот, интересно посмотреть, способны ли другие полимеразы с подобным набором исходных данных (Pol i и Pol лямбда) принимать участие в репарации одноцепочечных разрывов.
Четвертой ДНК-полимеразой в клетках человека, обладающей dRP-лиазной активностью, является митохондриальная ДНК-полимераза гамма. Pol гамма – единственная ДНК-полимераза, обнаруженная в митохондриях, следовательно, она ответственна за все преобразованиях ДНК, происходящие в этой органелле. Митохондрия является объектом интенсивного повреждения ДНК активными формами кислорода, генерируемыми во время окислительного фосфорилирования. Эти повреждения эффективно репарируются набором митохондриальных белков, которые включают в себя АP-эндонуклеазу, Pol гамма,
мтДНК-лигазу. Сам процесс репарации сходен с однонуклеотидным процессом BER в ядерной ДНК.
Таблица. ДНК-гликозилазы и эндонуклеазы клеток микроорганизмов и человека, участвующие в BER.
В универсальном механизме эксцизионной репарации как прокариоты, так и эукариоты гидролизуют 3–5-ю фосфодиэфирную связь с 3′-конца отповреждения. При этом прокариоты гидролизуют также 8-ю связьот 5’-конца измененного нуклеотида, тогда как у эукариотических организмовпроисходит одноцепочечный разрыв на расстоянии 21–25 нуклеотидов отповреждения со стороны его 5’-конца. Таким образом, прокариоты удаляютизмененный нуклеотид в составе 12–13-членных олигомеров, тогда как
эукариоты – в составе одноцепочечных фрагментов ДНК длиной в 27–29 нуклеотидов. Ферментная система, вносящая такие двойные одноцепочечные
разрывы, получила название эксцизионной нуклеазы (эксцинуклеазы). Образующаяся в молекуле репарируемой ДНК одноцепочечная брешь далее заполняется с помощью ДНК-полимеразы, а фосфодиэфирная связь в остающемся одноцепочечном разрыве восстанавливается ДНК-лигазой.
NER устраняет ДНК-повреждения посредством вырезания олигонуклеотида, содержащего это повреждение, с образованием бреши в ДНК размером ~ 30 нуклеотидов. Эта брешь застраивается одной или двумя полимеразами В семейства Pol дельта или Pol епсилон. Любая из этих ДНК-полимераз способна застраивать подобную брешь в реконструированной системе, содержащей очищенные белки млекопитающих, in vitro. При изучении NER в системе фибробластов человека с нарушенной проницаемостью и в ядерном экстракте HeLa клеток обнаружили, что обе полимеразы необходимы для восстановления ДНК после УФ-облучения. Исследования
в дрожжевых системах показали, что та или другая ДНК-полимераза существенно дополняют репарацию УФ-поврежденной ДНК. Более поздние исследования показали, что оба фермента необходимы для эффективной NER репарации в дрожжевых экстрактах. Pol дельта и Pol епсилон способны вести процессивный синтез ДНК при наличии дополнительного фактора PCNA, который «надевается» на ДНК с помощью пятисубъединичного комплекса RFC. Репарационный синтез как Pol дельта, так и Pol епсилон также требует присутствия этих факторов процессивности.
Эксцизионная репарация нуклеотидов: введение
ЭР — эксцизионная репарация ДНК; ЭР-комплекс включает ПАРП , XRCC1 , ДНК-лигазу III и ДНК-полимеразу бета .
Два пути осуществления NER:
1. Гидролиз фосфодиэфирной связи по 3′- или 5′- концу на некотором расстоянии от ошибочно спаренного (поврежденного) нуклеотида, который далее целиком удаляется под действием 5′->3′- (или 3′->5′-) экзонуклеазы, гидролизующей
цепь ДНК нуклеотид за нуклеотидом в соответствующем направлении от первоначального одноцепочечного разрыва в репарируемой ДНК. Образующаяся брешь далее заполняется ДНК-полимеразой. Такой механизм репарации реализуется у E. coli и человека для вырезания неповрежденных (немодифицированных) ошибочно спаренных нуклеотидов. Механизм последовательного эндо- и экзонуклеазного расщепления ДНК не используется для удаления поврежденных (измененных) нуклеотидов. Это связано с тем, что такие нуклеотиды часто являются ингибиторами экзонуклеаз.
2. Функционирует у всех исследованных видов организмов и заключается в использовании ферментной системы, которая вносит одноцепочечные разрывы по обе стороны от поврежденного нуклеотида на некотором расстоянии от него с последующим удалением одноцепочечного фрагмента ДНК, содержащего измененный нуклеотид.
Гидролизуется 3-5-фосфодиэфирную связь с 3′-конца от повреждения. При этом прокариоты гидролизуют также 8-связь от 5′-конца
измененного нуклеотида, тогда как у эукариотических организмов происходит одноцепочечный разрыв на расстоянии 21-25 нуклеотидов от повреждения со стороны его 5′-конца. Таким образом, прокариоты удаляют измененный нуклеотид в составе 12-13-членных олигомеров, тогда как эукариоты — в составе одноцепочечных фрагментов ДНК длиной в 27-29 нуклеотидов. Ферментная система, вносящая такие двойные одноцепочечные разрывы, получила название эксцизионной нуклеазы (эксцинуклеазы) . Образующаяся в молекуле репарируемой ДНК одноцепочечная брешь далее заполняется с помощью ДНК-полимеразы, а фосфодиэфирная связь в остающемся одноцепочечном разрыве восстанавливается ДНК-лигазой.
В отличие от ЭРО и прямых реверсий, которые специфичны к достаточно узкому кругу повреждений ДНК, система ЭРН, хотя и с разной эффективностью, удаляет все возможные повреждения, и потому роль ее в поддержании стабильности генома велика. ЭРН детально изучена у E. coli и активно изучается в клетках
дрожжей и человека, причем в последних благодаря раскрытию генетической природы таких заболеваний, как пигментная ксеродерма ( ХР ), синдром Кокейна ( СS ) и заболевания трихотиодистрофия ( ТТD)
В ЭРН задействовано 6-8 генов у E. coli и до 30 у человека.
Если повреждение в транскрибируемой (матричной) нити ДНК задерживает продвижение РНК-полимеразы, то специальный белковый фактор TRCF ( transcription-repair coupling factor ) сталкивает РНК-полимеразу и связывает с этим местом комплекс ферментов репарации. См. Сопряжение транскрипции и репарации ДНК у E/coli.
Ферментный ансамбль осуществляющий первые 3 стадии процесса NER называется эксцинуклеазой.
В системе эксцизионной репарации ДНК путем удаления нуклеотидов поврежденные азотистые основания вырезаются в составе олигонуклеотидов.
NER: регуляция
Для клеток животных не характерен SOS-ответ, свойственный клеткам E. coli и представляющий собой суммарную реакцию бактериальной клетки на повреждение ДНК различными агентами,
проявляющийся в усилении транскрипции генов NER. Посттрансляционные модификации белков репарации, происходящие в ответ на повреждение ДНК, не влияют на активность эксцинуклеазы человека.
Обнаружено, что повреждения ДНК стабилизирует белок р53- белок-супрессор опухолевого роста, являющийся регулятором транскрипции. Имеются данные о том, что белок р53 может взаимодействовать с белками XPB и RPA, необходимыми для NER. Однако клетки с инактивированными генами р53 (p53(-/-)), как и клетки дикого типа, эффективно удаляют из поврежденной ДНК два основных фотопродукта, возникающих под действием УФ-света, и обладают такой же устойчивостью к УФ. Поэтому считается, что белок р53 не оказывает прямого влияния на NER. Белки Cdk7 и циклин H, которые образуют Cdk-активирующую киназу, входят в состав комплекса TFIIH, что позволяет предполагать наличие связи репарации ДНК с фазами клеточного цикла.
ЭРН (NER): сопряжение с транскрипцией (ТЭРН)
Транскрибируемые последовательности нуклеотидов ДНК, особенно
в матричной цепи, репарируются с большей скоростью, чем нетранскрибируемые последовательности. В клетках больных с синдромом Кокайна не наблюдается такой асимметрии в репарации.
В клетках E. coli белковый фактор, кодируемый геном mfd и сопрягающий транскрипцию и репарацию, замещает остановившиеся перед повреждением молекулы РНК-полимеразы, что приводит к диссоциации транскрипционного комплекса. При этом он привлекает экзонуклеазный репаративный комплекс к поврежденному участку ДНК. См. Сопряжение транскрипции и репарации ДНК (ТЭРН) у E/coli
В клетках животных ген CSB кодирует белок с молекулярной массой 160 кДа, который содержит хеликазный домен и, возможно, выполняет те же функции, что и белок Mfd у E. coli . На основе поведения клеток с мутантными генами белков CSA и CSB разработана модель механизма, с помощью которого обеспечивается асимметричная репарация цепей ДНК. В соответствии с этой моделью РНК-полимераза II , остановившаяся в процессе транскрипции
перед поврежденным участком ДНК, распознается комплексом CSA-CSB и перемещается в сторону от повреждения без разрушения четвертичной структуры транскрипционного комплекса. Одновременно комплекс CSA-CSB привлекает компоненты репаративной системы XPA и TFIIH к месту повреждения ДНК и помогает сборке эксцинуклеазного комплекса. Нуклеотиды поврежденной цепи вырезаются, и брешь репарируется. После этого РНК-полимераза в составе транскрипционного комплекса продолжает транскрипцию.
NER: механизм
Процесс NER можно разделить на четыре этапа: а) распознавание поврежденного участка ДНК; б) двойное надрезание (инцизия) цепи ДНК по обеим сторонам поврежденного участка и его удаление (эксцизия); в) заполнение бреши в процессе репаративного синтеза; г) лигирование оставшегося одноцепочечного разрыва ДНК.
NER человека распознает и удаляет одиночные ошибочно спаренные нуклеотиды, а также петли длиной в 1-3 нуклеотида. NER человека способна различать цепи ДНК в случае распознавания поврежденных
нуклеотидов. Показано, что при наличии в ДНК димеров тимина циклобутанового типа вырезание нуклеотидов происходит исключительно из поврежденной цепи. Механизм такого распознавания в настоящее время неизвестен, как и молекулярный механизм узнавания самих поврежденных оснований. Система способна распознавать повреждения как сильно, так и слабо деформирующие вторичную структуру ДНК. При этом не обнаружена линейная зависимость между коэффициентом специфичности нуклеазы (kcat/km) и уровнем деформации двойной спирали ДНК. Показано, что в процессе распознавания участвуют белковые комплексы XPA/RPA, которые преимущественно связываются с поврежденной ДНК, и TFIIH, обладающий АTP-зависимой ДНК-расплетающей активностью. Последний взаимодействует с поврежденным участком ДНК и по аналогии с соответствующим механизмом у E. coli локально раскручивает ДНК, создавая основной преинцизионный комплекс с поврежденной ДНК.
Установлено, что три фермента репарации, обладающие узкой
субстратной специфичностью: ДНК-фотолиаза (удаление пиримидиновых димеров), урацилгликозилаза (удаление урацила из ДНК) и экзонуклеаза III (гидролиз ДНК в AP-сайтах), втягивают поврежденный участок из двойной спирали в полость фермента, что приводит кофактор или аминокислотные остатки активного центра этих ферментов в непосредственный контакт с расщепляемыми связями ДНК. Не исключено, что система эксцинуклеазы действует таким же образом.
Основные этапы функционирования NER, следующие за распознаванием поврежденного участка ДНК, представлены на рис. I.59. После связывания комплекса XPA-RPA с измененным участком ДНК, XPA взаимодействует с комплексом TFIIH, который создает преинцизионный комплекс, что сопряжено с гидролизом ATP. ATP-зависимое расплетание ДНК комплексом TFIIH подготавливает ее к взаимодействию с двумя XP-белками, обладающими нуклеазной активностью. XPG связывается с TFIIH и вносит одноцепочечный разрыв с 3′-конца повреждения. Аналогично комплекс ERCC1-XPF
взаимодействует с XPA в составе преинцизионного комплекса и способствует одноцепочечному разрыву с 5′-конца повреждения. Образование обоих разрывов является ATP-зависимым, и их расположение на ДНК высокоспецифично. Как правило, происходят разрывы 5-й и 24-й фосфодиэфирных связей соответственно от 3′- и 5′-концов поврежденных участков. Однако расположение точек разрывов может варьироваться.
Таким образом, в результате подобных одноцепочечных надрезов ДНК может освобождаться фрагмент длиной 24-32 нуклеотида с преобладанием фрагментов длиной 27-29 нуклеотидов. На расположение сайтов одноцепочечных разрывов влияют характер повреждения и последовательности нуклеотидов (контекст), окружающих поврежденный участок. Ту же самую картину инцизии обнаруживают in vivo в ооцитах Xenopus и у Schizosaccharomyces pombe. На этом основании делают вывод об универсальном механизме эксцизионной репарации у эукариот.
Репаративный синтез ДНК у человека является PCNA-зависимым,
т.е. может осуществляться с участием ДНК-полимераз Polдельта и Polэпсилон. PCNA связывается с системой праймер-матрица под действием фактора репликации RFC, откуда следует, что последний также участвует в репаративном синтезе ДНК. В опытах с бесклеточными системами моноклональные антитела к Polдельта специфически подавляют репаративный синтез. Однако оказалось, что в тех же высокоочищенных бесклеточных системах вместо Polдельта с аналогичным эффектом могут быть использованы Polэпсилон и даже фрагмент Кленова ДНК-полимеразы I E. coli. Это означает, что реконструированные из очищенных компонентов бесклеточные системы лишь в ограниченной степени имитируют биохимические процессы, происходящие в живых клетках. Считается, что обе ДНК-полимеразы — Polдельта и Polэпсилон участвуют в репаративном синтезе ДНК у человека.
ПАРП: модели управления процессом эксцизионной репарации ДНК
ПАРП — один из первых ядерных факторов, распознающих повреждение ДНК , и поэтому в идеальном случае управляет запуском механизма
репарации ДНК в живых клетках с места повреждения ДНК. Эта модель поддерживается идентификацией ЭР -комплекса, включающего ПАРП , XRCC1 , ДНК-лигазу III и ДНК-полимеразу бета . Присутствие ПАРП в таком мультипротеиновом комплексе доказывает, что этот фермент может направлять аппарат репарации ДНК к сайтам повреждения ДНК in vivo и облегчает осуществление репарации по этому пути.
XRCC1-белок действует как молекулярные «леса», формируя ЭР-комплекс путем индивидуального взаимодействия с каждым компонентом. С тех пор, как была обнаружена возможность поли(АДФ-рибозил)ирования XRCC1-белка in vitro, можно предположить, что ПАРП способен регулировать активность комплекса путем модифицирования XRCC1-белка in vivo и нарушать его способность взаимодействовать с другими компонентами комплекса. Было показано, что сверхэкспрессия XRCC1-белка подавляет активность ПАРП в живых клетках.
Аналогично ДНК-лигаза III ингибирует активность ПАРП in vitro, когда ее количества превышают количества ПАРП.
ПАРП может также рекрутировать факторы репарации ДНК путем модификации хроматиновых белков. Длинные цепи ПАР действительно способны направить ферменты репарации к сайтам разрывов ДНК значительно быстрее, нежели если они ищут повреждение сами по всему ядру. Такая модель согласуется с активностью ПАРП перед и/или после удаления поврежденных оснований.
NER: структура и функции белков
В табл. I.21 суммированы свойства белков животных, участвующих в NER. Большинство этих белков существует in vivo в виде комплексов, поэтому ферментативные активности, обнаруживаемые у отдельных белков в очищенном состоянии, могут не иметь прямого отношения к их функциям в системе NER.
XPA — белок с молекулярной массой 31 кДа, обладает доменом типа «цинковые пальцы», участвует в распознавании поврежденного участка ДНК, взаимодействует с другими компонентами системы и может функционировать в качестве фактора нуклеации для экзонуклеазы. XPA взаимодействует своим N-концевым доменом
с гетеродимером ERCC1-XPF, а С-концевым доменом — с TFIIH.
Белок RPA (HSSB) образует комплекс с XPA и усиливает его специфичность в отношении поврежденной ДНК. RPA (HSSB) — тример, состоящий из белковых субъединиц р70, р34 и р11, необходим для репликации ДНК и репаративного синтеза, а также для прохождения этапа двойного надреза ДНК во время эксцизионной репарации. Он обладает умеренным сродством к поврежденной ДНК.
TFIIH — олигомерный комплекс, в состав которого входят белки р89, р80, р62, р44, р41, р38 и р34. Этот белковый комплекс был открыт как один из семи основных факторов транскрипции, необходимых для эффективного функционирования РНК-полимеразы II . Субъединица р89 идентична белку репаративного комплекса XPB. Обнаружено отсутствие функциональной комплементации между бесклеточными экстрактами клеток с мутантными белками XPB и XPD, определяемой по восстановлению репарирующей активности в смешанных экстрактах. Комплекс TFIIH представляет собой
фактор репаративной системы. Белки XPB и XPD являются ДНК-зависимыми АТРазами, обладают хеликазными доменами и могут (как и сам фактор TFIIH) вызывать диссоциацию гибридов, образованных между короткими фрагментами ДНК и одноцепочечной ДНК.
XPC — белок с молекулярной массой около 125 кДа, существует в виде гетеродимера в комплексе с белком р58, который является гомологом белка Rad23 дрожжей (HHR23B). XPC слабо связывается с TFIIH и очень прочно — с одноцепочечной ДНК.
ERCC1/XPF — прочный белковый комплекс, с которым взаимодействует белок XPA, обладающий эндонуклеазной активностью, специфичной в отношении одноцепочечной ДНК.
XPG — белковый комплекс, обладающий эндонуклеазной активностью, специфичной в отношении одноцепочечной ДНК; вовлекается в эксцизионный комплекс посредством взаимодействия с TFIIH и RPA.
ЭРН (NER): генетика
Гены NER E. coli, uvrA, uvrB и uvrC не обнаруживают гомологии с соответствующими генами человека. Гены NER дрожжей и человека
высокогомологичны, и энзимология эксцизионной репарации также обладает большим сходством. По крайней мере, три заболевания у человека вызываются генетическими нарушениями системы эксцизионной репарации: пигментная ксеродерма, синдром Кокейна и трихотиодистрофия.
Кожа больных пигментной ксеродермой обладает повышенной чувствительностью к дневному свету, что проявляется в виде фотодерматозов, включая рак кожи. В ряде случаев отмечены аномалии нервной системы, причиной которых являются мутации в одном из семи генов: XPA, XPB, XPC, XPD, XPE, XPF, XPG. Однако описаны больные с классическими симптомами пигментной ксеродермы, но с ненарушенной системой NER. Для клеток этих больных характерны изменения в так называемой пострепликативной репарации .
Больным с синдромом Кокейна присущи нарушения роста, умственная отсталость, катаракты, повышенная чувствительность к свету с сопутствующими дерматозами. Обнаружены мутации в двух группах генов, приводящие к этому заболеванию. У больных
с мутантными генами CS-A или CS-B клетки способны нормально репарировать УФ-повреждения ДНК. У другой группы больных обнаружены мутации в генах XPB, XPD или XPG.
У больных трихотиодистрофией со смешанными симптомами выявлены мутации в генах XPB или XPD. Классические симптомы этого заболевания, по-видимому, являются следствием мутации в гене транскрипционного фактора TFIIH.
Получение мутантов с измененной NER у грызунов позволило разбить такие гены на 11 групп комплементации, большинство из которых соответствует группам комплементации XP и CS человека. Часть соответствующих генов человека удалось клонировать, используя их способность исправлять (комплементировать) генетические дефекты в культивируемых мутантных клетках грызунов. Эти гены получили название кросс-комплементирующих генов эксцизионной репарации ( ERCC — excision repair cross complementing ). Среди них гены XPE и ERCC6-ERCC11 не требовались для прохождения основных реакций эксцизионной репарации, и их функция неизвестна.
Эксцизионная репарация нуклеотидов (ЭРН) E. coli
У E. coli эксцинуклеаза формируется, в результате димеризации двух молекул белка UvrА в присутствии АТР и связывания с одной молекулой UvrB. Гетеротример UvrA2B связывается с ДНК и с помощью 5′-3′-ДНК-геликазной активности перемещается вдоль ДНК в поисках повреждения. Такова предполагаемая картина узнавания эксцинуклеазой UvrAВС неспецифического повреждения в ДНК [ Hoeijmakers J., 1993 ]. Результатом узнавания повреждения этим комплексом явится внедрение субъединицы UvrB в ДНК, сопровождающееся конформационными изменениями ДНК в сайте внедрения ( изломы , локальная денатурация), диссоциация обеих субъединиц UvrА из комплекса и последующее связывание с ним молекул UvrD и UvrC. Гетеродимер UvrBC из состава нового комплекса делает два разрыва в поврежденной нити ДНК на расстоянии 8 н. с 5′-конца от повреждения и 5 н. с 3′-конца, катализируемых субъединицами UvrC и UvrB соответственно, что приводит к появлению 12-13-мерного олигонуклеотида,
вытесняемого из комплекса с помощью геликазы UvrD. Образующаяся при этом брешь заполняется ДНК-полимеразой I, синтезирующей ДНК по неповрежденной матрице, и сшивается ДНК-лигазой. Описанный процесс зависит от АТР. Действительно, АТР стимулирует димеризацию молекул UvrA, гидролиз АТР необходим для образования комплекса UvrA2B и проявления его геликазной активности, АТР необходим для формирования комплекса UvrB-ДНК и, наконец, для описанной выше бимодальной инцизии ДНК [ Friedberg E.C., Walker G.C., ea., 1991 ]. У E. coli транскрипционно-зависимую ветвь ЭРН (ТЭРН) осуществляет продукт гена mfd (mutation frequency decline). Этот ген был открыт за 35 лет до того, как его продукт признали фактором TRCF. Последний способствует диссоциации РНК-полимеразы от дефектной транскрибируемой нити ДНК, и связывается с субъединицей UvrA эксцинуклеазного комплекса. Иными словами, именно белок Mfd отыскивает повреждения на транскрибируемой нити и направляет ТЭРН на эту мишень.
Согласно недавним наблюдениям, два главных белка системы ДКНО — MutL и MutS — необходимы для ТЭРН [106 Melon I., Champl G.N., 1996106]. Причина такой взаимосвязи систем ДКНО и ТЭРН или заимствования белков MutL и MutS для выполнения сходных функций пока остается загадочной, хотя сам факт несомненен и нашел подтверждение также и для белков Msh2, Mlh1, Pms1 и Msh3 у S. cerevisiae [ Sweder K.S., ea, 1996 ].
Эксцизионная репарация нуклеотидов (ЭРН) у человека
Большому прогрессу в раскрытии механизма ЭРН у эукариот мы обязаны наследственному заболеванию человека — пигментной ксеродерме .
Изучение молекулярных основ этого заболевания выявили их сопряженность с дефектами системы ЭРН, результатом чего и явилось раскрытие генетического контроля ЭРН. Комплементационный анализ различных клеточных линий ХР определил 8 комплементационных групп (7 от XPA до XPG и одна «вариантная форма» XPV) [ Friedberg E.C., Walker G.C., ea., 1991 ]), а комплементационная
коррекция возможных мутантных линий клеток китайского хомячка, чувствительных к УФ-свету, способствовала выявлению дополнительных генов системы ЭРН. Последние получили название гены ERCC (excision repair cross-complimenting), и поэтому в названии генов и белков ЭРН используются обе аббревиатуры. Хотя детали процесса ЭРН у человека не ясны, так как не определена роль целого ряда генов, в первом приближении он выглядит так ( табл. 3 ) [ Lehmann A.R., 1995 , Lindahl T., ea, 1997 , Hoeijmakers J., 1993 ]. Белок ХРА в комплексе с онДНК-связующим белком RPA ( репликационным белком А ), транслоцируясь вдоль онДНК, опознает конформационнoе повреждениe. Взаимодействуя через другой участок белка ХРА с базальным фактором транскрипции TFIIH (две из субъединиц которого, белки ХРВ и ХРD , обладают геликазной активностью с противоположной ориентацией раскручивания днДНК), они образуют комплекс, который расплетает ДНК вокруг повреждения (в состав этого комплекса входит и белок ХРС
с неясными функциями). Через третий участок белка ХРА к комплексу примыкает гетеродимер ERCC1-XPF , который вносит однонитевой разрыв в ДНК с 5′-конца на расстоянии 16-25 н. от повреждения, тогда как белок XPG , входящий в комплекс белков эксцинуклеазы через взаимодействие с белком RPA, делает надрез с 3′-конца на расстоянии 2-9 н. (в разрезании принимает участие и белок ХРС , а белок ХРЕ активирует реакцию). В результате бимодальной инцизии участок ДНК размером около 29 н. высвобождается, а образующаяся брешь ресинтезируется с помощью ДНК-полимеразы епсилон или ДНК-полимеразы дельта , сопутствующего репликации фактора PCNА , репликационного фактора C-RFС и ДНК-лигазы I .
Представленная картина во многом основана на реконструировании процесса в открытой системе с участием 10 хорошо очищенных белков [ Wood R.D., 1994 ]. Как видно, эукариотическая система ЭРН лишь функционально подобна прокариотической. В ней задействовано значительно больше белков, число которых должно еще возрасти
за счет белков, осуществляющих разборку и сборку хроматина. Раскрытие природы ТЭРН у человека также связывают с пониманием молекулярного дефекта,приводящего к развитию двух мультисистемных генетических заболеваний — синдрома Кокейна (СS) и трихотиодистрофии ( ТТD ). При обоих заболеваниях соматические клетки больных оказались неспособны к ТЭРН. Классические случаи CS оказались связанными с повреждениями в двух генах, CSA и CSB, а редкие смешанные формы CS+XP (CS с дополнительными симптомами ХР), с повреждениями в генах ХРВ, ХРD и ХРG. При ТТD дефекты были обнаружены также в генах XPB и XPD и в новом пока не клонированном гене ТТDА, продукт которого является частью корового домена транскрипционного фактора TFIIH [ Lehmann A.R., 1995 , Lindahl T., ea, 1997 ].
Фактор TFIIH состоит из 6 субъединиц, две из которых — белки XPB и XPD. Связываясь с белками РНК-полимеразного комплекса, кор TFIIH принимает участие в инициировании транскрипции [ Buratowski S., 1994 ], a вкупе с белками репарационного
комплекса кор этого фактора участвует в ЭРН. Если допустить, что белки CSA и CSB способствуют превращению транскрипционного комплекса в репарационнный [ Lindahl T., ea, 1997 , Bregman D.B., 1996 ], то следствием повреждения этих белков может стать дефектность в ТЭРН. Таким образом, либо прямыми воздействиями на коровый домен фактора TFIIH (мутации в генах XPB, XPD и TTDA), либо косвенными (через гены CSA и CSB) можно модифицировать способность этого фактора к диссоциации из РНК- полимеразного комплекса для участия в ТЭРН. Что касается белка XPG, то он, подобно своему гомологу из S. cereivisiae — белку Rad2 [ Bregman D.B., 1996 ], способен взаимодействовать c TFIIH, что допускает возможность его воздействия на TFIIH для участия в ТЭРН. Таковы гипотетические объяснения взаимосвязи молекулярных дефектов при CS и ТTD с нарушениями ТЭРН, которые могут быть полезными для раскрытия механизма ТЭРН у эукариот. Связь же между молекулярными дефектами и клиническими проявлениями
рассматриваемых наследственных заболеваний пока не имеет даже упрощенных толкований [ Kolodner R., 1995 ].
Так случилось, что изучение молекулярных механизмов ЭРН и ТЭРН на клетках человека развивались параллельно, или даже опережая исследования на популярной эукариотической модели дрожжей S. cerevisiae. В табл. 3 представлены известные функциональные аналоги системы ЭРН у дрожжей. Несомненно, однако, что при исследовании детального механизма ЭРН именно эта модельная система будет играть ведущую роль. Например, немаловажной особенностью системы ЭРН у E. coli является ее связь с SOS-функциями клетки. Действительно, центральные гены системы ЭРН, uvrA и uvrB, находятся под контролем SOS-регулона [ Friedberg E.C., Walker G.C., ea., 1991 ]. Системы, подобной SOS, у эукариот пока не обнаружено. Однако целый ряд генов системы ЭРН у дрожжей, таких, как RAD2, RAD7, RAD23, CDC8 и CDC9, индуцируется при повреждении ДНК, а последние два гена дополнительно регулируются клеточным циклом [ Friedberg
E.C., Walker G.C., ea., 1991 ].
Природу этой индукции еще предстоит выяснить.
ЧАСТЬ III. ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ
25. МЕТАБОЛИЗМ ДНК
25.2. Репарация ДНК
Большинство клеток содержит только одинарный или двойной набор геномной ДНК. Поврежденные белки и молекулы РНК могут быстро заменяться новыми благодаря информации, закодированной в ДНК. но сами молекулы ДНК заменяться не могут. Поддержание целостности информации в ДНК — важнейшая задача клетки, осуществляемая с помощью сложного набора систем репарации ДНК. ДНК может разрушиться в результате разных процессов; некоторые из них спонтанны, другие катализируются факторами внешней среды (см. гл. 8 в т. 1). Даже сам процесс репликации может иногда искажать содержащуюся в ДНК информацию, если в результате ошибки образуются неправильные пары оснований (например, пара G-T).
Химия повреждений ДНК разнообразна и сложна. Клеточный ответ на конкретное повреждение связан с активностью широкого круга ферментных систем, катализирующих одни из наиболее интересных химических трансформаций в метаболизме ДНК. Сначала мы рассмотрим последствия изменений в последовательности ДНК, а затем специфические системы репарации.
Онкологические заболевания связаны с мутациями
Лучший способ проиллюстрировать значение репарации ДНК — рассмотреть последствия неисправленных повреждений. Наиболее опасны такие изменения ДНК, которые в результате репликации и передачи новым поколениям клеток становятся постоянными. Стабильное изменение нуклеотидной последовательности ДНК называется мутацией. Мутации могут выражаться в замене одной пары оснований на другую (однонуклеотидные замены) или же происходит вставка и выпадение одной или нескольких пар оснований (инсерции и делеции). Если мутация затрагивает несущественный участок ДНК или не имеет принципиального значения для работы гена, ее называют молчащей мутацией. Изредка мутации обеспечивают своему хозяину биологическое преимущество. Однако в большинстве случаев проявляющиеся мутации являются нейтральными или вредоносными.
У млекопитающих наблюдается четкая корреляция между накоплением мутаций и раком. В ходе простого теста, разработанного Брюсом Эймсом, можно выявить способность химического соединения вызывать определенные легко выявляемые мутации в специальных бактериальных штаммах (рис. 25-21). Согласно этому тесту, совсем немногие вещества, которые мы используем повседневно, являются мутагенами. Однако более 90% соединений, в экспериментальных условиях, оказывающих канцерогенное действие на животных, в тесте Эймса проявляют мутагенную активность. Учитывая четкую корреляцию между мутагенезом и канцерогенезом, тест Эймса для бактериальных мутагенов широко применяется в качестве быстрого и дешевого метода выявления канцерогенов человека.
Рис. 25-21 Тест Эймса для выявления канцерогенов, основанный на мутагенности. Штамм Salmonella typhimurium, несущий мутацию, инактивирующую фермент метаболического пути биосинтеза гистидина, высевают на среду без гистидина. Растут только единичные клетки. а — несколько мелких колоний S. typhimurium, которые, тем не менее, вырастают на не содержащей гистидин среде. несут спонтанные обратные мутации, восстанавливающие биосинтез гистидина. Три чашки б, в и г с такой средой засевают равным числом клеток. Затем в центр каждой чашки помешают диски фильтровальной бумаги с разной концентрацией мутагена. Мутаген значительно увеличивает вероятность возникновения обратной мутации и, соответственно, число выросших клеток в колонии. Чистая зона вокруг фильтровальной бумаги показывает, что концентрация мутагена здесь настолько высокая, что оказывается летальной для клеток. По мере того как мутаген диффундирует, удаляясь от диска фильтровальной бумаги, его концентрация понижается до сублетальных концентраций, которые индуцируют обратные мутации. Мутагены можно сравнивать на основании их влияния на частоту возникновения мутаций. Поскольку после проникновения в клетку многие соединения претерпевают разнообразные химические превращения, соединения иногда тестируют на мутагенность после их предварительной инкубации с экстрактом печени. Мутагенность некоторых веществ проявляется только после такой обработки.

В геноме типичной клетки млекопитающего за 24 ч аккумулируется много тысяч повреждений. Однако в результате репарации ДНК менее одного повреждения из 1000 становится мутацией. Молекула ДНК относительно стабильна, но при отсутствии систем репарации кумулятивный эффект многих нечастых, но разрушительных реакций может привести к гибели организма. ■
Все клетки имеют несколько систем репарации ДНК
Количество и разнообразие систем репарации отражает важность репарации ДНК для выживания клетки, а также многообразие способов повреждения ДНК (табл. 25-5). Некоторые распространенные типы повреждений, например, пиримидиновые димеры (см. рис. 8-31), могут устраняться с помощью нескольких систем репарации. Многие процессы репарации ДНК очень неэффективны энергетически в отличие от большинства метаболических путей, в которых обычно каждая молекула АТР на счету и используется оптимальным образом. Но когда на карту поставлена целостность генетической информации, энергетические затраты не имеют значения.
Репарация ДНК возможна в значительной степени благодаря тому, что молекула ДНК состоит из двух комплементарных цепей. Повреждение ДНК в одной цепи может быть удалено и аккуратно исправлено благодаря использованию неповрежденной комплементарной цепи в качестве матрицы. Далее мы рассмотрим основные типы систем репарации, начиная с репарации редких неправильно спаренных нуклеотидов, остающихся после репликации.
Репарация ошибочно спаренных оснований.
Коррекция редких ошибочно спаренных оснований, остающихся после репликации в Е. coli, увеличивает общую точность репликации еще на 2-3 порядка. Неправильно спаренные основания почти всегда корректируются в соответствии с информацией в старой (матричной) цепи, поэтому система репарации должна каким-то образом различать исходную и заново синтезированную цепи. Для этого клетка метит матричную ДНК метальными группами. Система репарации ошибочно спаренных оснований Е. coli включает по меньшей мере 12 белковых компонентов (табл. 25-5), которые либо дискриминируют цепи, либо участвуют в самом процессе репарации.
Таблица 25-5. Типы систем репарации ДНК у E. coli

Механизм дискриминации цепей не установлен для большинства бактерий и эукариот, но хорошо изучен у Е. coli и некоторых родственных бактерий. У этих прокариот дискриминация цепей основана на действии Dam-метилазы, которая, как вы помните, метилирует ДНК в положении N6 остатка аденина в составе последовательностей (5’) GATC. Сразу после прохождения репликативной вилки наступает короткий период (несколько секунд или минут), когда матричная цепь метилируется, а вновь синтезированная цепь нет (рис. 25-22). Временно неметилированные последовательности GATC во вновь синтезированной цепи позволяют отличить новую цепь от матричной. Ошибочно встроенные основания вблизи наполовину метилированной последовательности GATC устраняются исходя из информации, содержащейся в метилированной родительской (матричной) цепи. Исследования in vitro показали, что, если последовательности GATC метилированы в обеих цепях, исправляется небольшое число ошибок; если ни одна из цепей не метилирована, репарация происходит, но ни одна из цепей не имеет преимуществ. Клеточные системы репарации, основанные на метилировании, аффективно устраняют однонуклеотидные замены на расстоянии до 1000 п. н. от наполовину метилированной последовательности GATC.
Рис. 25-22. Метилирование и репарация ошибочно спаренных оснований. Метилирование цепей ДНК у Е. coli может служить для дискриминации родительских (матричных) цепей и вновь синтезированных цепей, что имеет решающее значение для репарации однонуклеотидных замен; см. рис. 25-23. Метилирование происходит по положению N6 остатков аденина в последовательностях 5’(GATC). Эта последовательность представляет собой палиндром (см. рис. 8-18 в т. 1), расположенный на двух цепях в противоположных ориентациях.

Как такие относительно удаленные последовательности GATC направляют коррекцию ошибочно спаренных оснований? Механизм показан на рис. 25-23. Белок MutL образует комплекс с белком MutS, и этот комплекс связывается со всеми аномальными парами оснований (кроме С-С). Белок MutH связывается с MutL и последовательностями GATC, с которыми встретился комплекс MutL-MutS. ДНК с обеих сторон от аномальной пары протягивается через комплекс MutL-MutS, создавая петлю. Одновременное продвижение обоих концов петли через комплекс равнозначно движению комплекса вдоль ДНК одновременно в двух направлениях. Белок MutH обладает сайт-специфической эндонуклеазной активностью, которая не проявляется ло тех пор. пока комплекс не встречает наполовину метилированную последовательность GATC. В этом месте MutH катализирует расщепление неметилированной цепи на 5’-конце остатка G в составе последовательности GATC, что помечает цепь для последующей репарации. Дальнейшие стадии метаболического пути зависят от того, где именно располагается ошибочно спаренное основание относительно места расщепления (рис. 25-24).
Рис. 25-23. Модель ранних стадий репарации, основанной на метилировании ДНК. Участвующие в данном процессе белки Е. coli (см. табл. 25-5) были выделены и очищены. Узнавание последовательности 5′(GATC) и распознавание ошибочно спаренного основания — специфические функции белков MutL и MutS соответственно. Белок MutH образует комплекс с MutS в точке аномального спаривания. ДНК проходит через этот комплекс таким образом, что комплекс движется одновременно в двух направлениях вдоль ДНК до тех пор, пока не придет в соприкосновение с белком MutH, связанным с полумети- лированной последовательностью GATC. Белок MutH расщепляет неметилированную цепь по остатку гуанина в этой последовательности со стороны 5′-конца. Затем комплекс, включающий ДНК-хеликазу II и одну из экзонуклеаз, расщепляет неметилированную цепь от этой точки до ошибочно спаренного основания (см. рис. 25-24).

Рис. 25-24. Завершение репарации, основанной на метилировании ДНК. Действуя совместно, ДНК-хеликаза II, SSB и одна из четырех экзонуклеаз удаляют участок новой цепи между местом расщепления белком MutH и точкой, расположенной сразу за ошибочно спаренным основанием. Выбор экзонуклеазы зависит от расположения места расщепления относительно аномальной пары оснований. На схеме показаны альтернативные варианты. Образующуюся брешь заполняет ДНК-полимераза III (пунктирная линия), а ник ликвидирует ДНК-лигаза (не показано).

Если ошибочно спаренное основание находится со стороны 5′-конца от места расщепления (рис. 25-24, справа), неметилированная цепь раскручивается и разрушается в направлении 3′ —> 5′ от места расщепления, захватывая ошибочно спаренное основание, и этот участок замещается новой ДНК. Данный процесс требует совместного действия ДНК-хеликазы II, SSB, экзонуклеазы I или экзонуклеазы X (обе разрушают цепи ДНК в направлении 3′ —> 5′), ДНК-полимеразы III и ДНК-лигазы. Метаболический путь репарации ошибочно спаренных оснований, находящихся со стороны 3′-конца от точки расщепления, отличается лишь тем, что в нем участвует экзонуклеаза VII (которая расщепляет одноцепочечную ДНК в направлении 5′ —> 3′ или 3′ —> 5′) или RecJ-нуклеаза (которая расщепляет одноцепочечную ДНК в направлении 5′ —> 3′).
При оценке энергетической стоимости репарации однонуклеотидных замен у Е. coli выясняется, что это очень затратный процесс. Аномальная пара оснований может находиться на расстоянии 1000 и. н. и более от GATC. Расщепление и восстановление сегмента такой длины требует огромного количества активированных предшественников дезоксинуклеотидов для репарации одной нуклеотидной замены. Это еще раз подчеркивает значение целостности генома для клетки.
Все эукариотические клетки содержат несколько белков, структурно и функционально аналогичных бактериальным белкам MutS и MutL (но не MutH). У человека изменения в генах, кодирующих белки этого типа, вызывают некоторые наиболее распространенные наследуемые синдромы, связанные с предрасположенностью к онкологическим заболеваниям (доп. 25-1), что еще раз демонстрирует значение систем репарации для организма. У большинства эукариот, от дрожжей до человека.
основные гомологи MutS — белки MSH2 (от MutS homolog 2), MSH3 и MSH6. Гетеродимеры MSH2 и MSH6 обычно связываются с одиночными аномальными парами оснований, но хуже связываются с чуть более длинными неправильно спаренными петлями. У многих организмов более длинные участки аномального спаривания (от 2 до 6 п. н.) связываются с гетеродимерами MSH2/MSH3 или с гетеродимерами двух типов в тандеме. Гомологи MutL, особенно гетеродимер MLH1 и PMS1 (от. postmeiotic segregation), соединяются с комплексами MSH и стабилизируют их. Детальные исследования процесса репарации ошибочно спаренных оснований у эукариот активно продолжаются. В частности, пока неизвестен механизм, с помощью которого идентифицируются вновь синтезированные цепи ДНК, хотя было установлено, что последовательности GATC в этомучастия не принимают.
Эксцизионная репарация оснований.
Каждая клетка содержит класс ферментов, называемых ДНК -гликозилазами; они распознают наиболее распространенные повреждения ДНК (например, продукты дезаминирования цитозина и аденина; см. рис. 8-30. а) и удаляют поврежденные основания посредством расщепления N-гликозидной связи. Такое вырезание основания приводит к возникновению апуринового или апиримидинового сайта в ДНК, который называют АР-сайтом или сайтом, лишенным основания. Обычно каждая ДНК-гликозилаза специфична к одному типу нарушений.
Так, урацил-ДНК-гликозилаза, обнаруженная в большинстве клеток, специфически удаляет из ДНК урацил, который образуется при спонтанном дезаминировании цитозина. В мутантных клетках, утративших этот фермент, повышается доля мутаций в парах G = C по сравнению с мутациями в парах А = Т. Эта гликозилаза не удаляет
остатки урацила из РНК или остатки тимина из ДНК. Необходимость дискриминации тимина и урацила (продукта дезаминирования цитозина) для селективной репарации цитозина может быть одной из причин, по которой в состав ДНК входит тимин, а не урацил (см. с. 416 в т. 1).
У большинства бактерий есть только один тип урацил-ДНК-гликозилазы, а у человека существует по меньшей мере четыре типа с различной специфичностью — это показатель важности удаления урацила из ДНК. Наиболее распространенная урацил-гликозилаза человека, UNG, связывается с реплисомой и вырезает остатки урацила, случайно встроенные вместо тимина в ходе репликации. Дезаминирование остатков цитозина происходит в 100 раз быстрее в одноцепочечной ДНК, чем в двухцепочечной, и у человека есть специальный фермент hSMUG1, который удаляет любые остатки урацила в одноцепочечной ДНК в ходе репликации или транскрипции. Две другие ДНК- гликозилазы человека, TDG и MBD4, удаляют спаренные с гуанином остатки урацила и тимина, образованные дезаминированием цитозина или 5-метилцитозина соответственно.
Другие ДНК-гликозилазы распознают и удаляют различные поврежденные основания, включая формамидопиримидин и 8-гидроксигуанин (оба образуются при окислении пурина), гипоксантин (образуется при дезаминировании аденина) и алкилированные основания, такие как 3-метиладенин и 7-метилгуанин. В некоторых классах организмов также были идентифицированы гликозилазы, которые распознают другие дефекты, включая пиримидиновые димеры. Вспомните, что АР-сайты образуются также в результате медленного спонтанного гидролиза N-гликозидных связей в ДНК (см. рис. 8-30, б в т. 1).
Образованный ДНК-гликозилазой АР-сайт устраняют другие ферменты. Репарация происходит не просто путем присоединения нового основания и восстановления N-гликозидной связи. Вместо этого удаляется оставшийся дезокси- рибозо-5′-фосфат и встраивается новый нуклеотид. Этот процесс начинает одна из АР-эндонуклеаз, расщепляющих цепь ДНК, в которой содержится АР-сайт. Положение разреза относительно АР-сайта (со стороны 3′- или 5′-конца) зависит от типа АР-эндонуклеазы. Затем участок ДНК. включающий АР-сайт, удаляется, ДНК-полимераза 1 достраивает ДНК, а ДНК-лигаза ликвидирует оставшийся ник (рис. 25-25). У эукариот замену нуклеотидов выполняют специализированные полимеразы, как описано ниже.
Рис. 25-25. Эксцизионная репарация оснований. ① ДНК-гликозилаза распознает поврежденное основание и отщепляет его от дезоксирибозы в цепи ДНК. ② АР- эндонуклеаза расщепляет фосфодиэфирную связь около АР-сайта. ③ ДНК-полимераза I инициирует репарационный синтез, начиная от свободной 3′-гидроксильной группы в нике, при этом удаляется (с помощью 5′ —> 3′-экзонуклазной активности) часть поврежденной цепи и происходит замена ее на новую ДНК. ④ Ник, оставшийся после диссоциации ДНК-полимеразы I, ликвидирует ДНК-лигаза.

Эксцизионная репарация нуклеотидов.
Повреждения ДНК, которые вызывают значительные искажения спиральной структуры, обычно восстанавливаются системой эксцизионной репарации нуклеотидов. Этот путь репарации принципиально важен для выживания всех свободноживущих организмов. В этой системе репарации (рис. 25-26) мультисубъединичный фермент гидролизует две фосфодиэфирные связи по одной с каждой стороны от дефекта. У Е. coli и других бактерий ферментная система гидролизует 5-фосфодиэфирную связь с 3′-конца и 8-фосфодиэфирную связь с 5′-конца, создавая фрагмент из 12-13 нуклеотидов (в зависимости от того, одно или два основания повреждено). У человека и других эукариот ферментная система гидролизует 6-ю фосфодиэфирную связь с 3′-конца и 22-ю фосфодиэфирную связь с 5′-конца, образуя фрагмент из 27-29 нуклеотидов. В результате двустороннего разреза из дуплекса высвобождается нуклеотид с поврежденным участком, а образующаяся брешь заполняется у Е. coli ДНК-полимеразой I, а у человека — ДНК-полимеразой ε. Ник запечатывает ДНК-лигаза.
Рис. 25-26. Эксцизионная репарация нуклеотидов у Е. coli и человека. Основной путь репарации путем вырезания нуклеотидов почти одинаковый у всех организмов. ① Эксцизионная нуклеаза связывается с ДНК в месте крупного повреждения и расщепляет поврежденную цепь ДНК с обеих сторон от повреждения. ② Фрагмент ДНК длиной 13 нуклеотидов (13-мер) или 29 нуклеотидов (29-мер) удаляется с помощью хеликазы. ③ Брешь заполняется ДНК-полимеразой, а ④ оставшийся ник ликвидируется ДНК-лигазой.

У Е. coli ключевую роль в этом процессе исполняет эксцизионная нуклеаза АВС, состоящая из трех субъединиц: UvrA (Мr = 104 000), UvrB (Мr = 78 000) и UvrC (Мr = 68 000). Определение «эксцизионная нуклеаза» описывает уникальную способность этого ферментного комплекса катализировать двойное специфическое расщепление, что отличает его от обычных эндонуклеаз. Комплекс белков UvrA и UvrB (А2В) сканирует ДНК и связывается в месте повреждения. Затем димер UvrA диссоциирует, оставляя прочный комплекс UvrB — ДНК. После этого с UvrB связывается белок UvrC, и UvrB производит расщепление 5-й фосфодиэфирной связи со стороны 3’-конца от повреждения. Далее следует UvrC-опосредованное расщепление 8-й фосфодиэфирной связи со стороны 5′-конца. Образующийся фрагмент из 12-13 нуклеотидов удаляется ДНК-хеликазой. Получившаяся небольшая брешь устраняется ДНК-полимеразой I и ДНК-лигазой. Это основной путь репарации для многих типов повреждений (см. рис. 25-26. слева), включая циклобутановые пиримидиновые димеры, 6,4-фотоиродукты (см. рис. 8-31 в т. 1) и некоторые другие аддукты, включая бензо[а]пиренгуанин, который образуется в ДНК под воздействием табачного дыма. Нуклеолитическая активность эксцизионной нуклеазы АВС отличается от активности других эндонуклеаз тем, что она одновременно производит два разреза в ДНК.
Механизм работы эукариотических эксцизи- онных нуклеаз чрезвычайно схож с аналогичным механизмом бактериального фермента, хотя для двойной эксцизии в клетках эукариот необходимы 16 полипептидов, не имеющих сходства с субъединицами эксцизионной нуклеазы Е. coli. Как описано в гл. 26, эксцизионная репарация нуклеотидов и оснований у эукариот тесно связана с транскрипцией. Генетические дефекты системы эксцизионной репарации нуклеотидов — причина различных серьезных заболеваний человека (см. доп. 25-1).
Прямая репарация.
Некоторые типы повреждений устраняются без удаления нуклеотида или основания. Наиболее характерный пример прямая фотореактивация циклобутановых пиримидиновых димеров, которая осуществляется ДНК-фотолиазами. Пиримидиновые димеры образуются в индуцируемой УФ-облучением реакции, и фотолиазы используют энергию поглощенного света для устранения повреждения (рис. 25-27). Фотолиазы обычно содержат два кофактора, которые действуют как светопоглощающие агенты (хромофоры). Одним из хромофоров всегда является FADH. У Е. coli и дрожжей другой хромофор — фолат. В ходе реакции образуются свободные радикалы. В клетках плацентарных млекопитающих (в т. ч. у человека) ДНК-фотолиазы отсутствуют.
Рис. 25-27. Механизм реакции. Устранение пиримидиновых димеров фотолиазой. Энергия поглощенного света используется для ликвидации последствий фотореакции, вызвавшей повреждение. У Е. coli совместно работают два хромофора фотолиазы (Mr = 54 000): N5, N10-метенилтетрагидрофолилполиглутамат (MTHFpolyGlu) и FADH—. MTHFpolyGlu выполняет функцию антенны, поглощая фотоны синего света (300-500 нм). Энергия возбуждения передается на FADH—, и возбужденный флавин (*FADH—) отдает электрон пиримидиновому димеру, устраняя повреждение.

Другим примером может служить репарация нуклеотидов, дефект в которых вызван алкилированием. Модифицированный нуклеотид О6-метилгуанин образуется в присутствии алкилирующих агентов — это широко распространенное и сильно мутагенное повреждение (см. с. 419 в т. 1). В ходе репликации он легче образует пару с тимином, чем с цитозином, и в результате происходят замена пары G = C на А = Т (рис. 25-28). Прямую репарацию О6— метилгуанина производит О6-метилгуанин- ДНК-метилтрансфераза, которая катализирует перенос метальной группы на один из своих остатков Cys. Эта метилтрансфераза не истинный фермент, поскольку в результате однократного переноса метальной группы она остается метилированной и выбывает из процесса. Расход целой белковой молекулы для исправления одного поврежденного основания — очередная яркая иллюстрация приоритета целостности клеточной ДНК.

Поиному, но тоже прямому механизму идет репарация 1 -метиладенина и 3-метилцитозина. Аминогруппы остатков А и С иногда метилируются (обычно в одноцепочечной ДНК), и это влияет на правильность спаривания оснований. У Е. coli окислительное деметилирование подобных алкилированных нуклеотидов опосредует белок AlkB — представитель суперсемейства α-кетоглутарат- Fе2+ — зависимых диоксигеназ (рис. 25-29). (Описание другого фермента из этой группы см. в доп. 4-3 в т. 1.)
Рис. 25-28. Пример образования мутаций в результате повреждения ДНК. а — продукт метилирования O6— метилгуанин легче образует пару с тимином, чем с цитозином. б — при отсутствии репарации это приводит к мутации: после репликации G = C заменяется на А = Т.

Рис. 25-29. Прямая репарация алкилированных оснований белком AlkB. Белок AlkB — α-кетоглутарат-Fе2+-зависимая диоксигеназа; катализирует окислительное деметилирование остатков 1-метиладенина и 3-метил- цитозина.

Взаимодействие репликативных вилок с повреждением в ДНК может запустить подверженный ошибкам синтез ДНК через повреждение
Рассмотренные выше пути репарации обычно действуют в случае повреждений в двухцепочечной ДНК, при этом точность восстановления генетической информации поврежденной цепи до первоначального состояния обеспечивает неповрежденная цепь. Однако при некоторых типах повреждений, например, при двунитевых разрывах, поперечных сшивках или повреждениях в одноцепочечной ДНК, комплементарная цепь также оказывается разрушенной или отсутствует. Двунитевые разрывы и повреждения в одноцепочечной ДНК чаще всего возникают в тех случаях, когда репликативная вилка наталкивается на нерепарированное повреждение ДНК (рис. 25-30). Такие повреждения и поперечные сшивки ДНК, кроме того, могут быть результатом воздействия ионизирующей радиации и окислительных реакций.
Рис. 25-30. Повреждения ДНК и их влияние на репликацию. Если репликативная вилка наталкивается на неисправленное повреждение или разорванную цепь, репликация обычно останавливается. Слева: повреждение, оставшееся в нереплицированном одноцепочечном сегменте ДНК; справа: двухцепочечный разрыв цепи. В любом случае повреждение в одной цепи не может быть устранено ранее описанными механизмами, поскольку комплементарная цепь, необходимая для точной репарации, повреждена или отсутствует. В таких случаях есть два возможных пути репарации: рекомбинационная репарация ДНК (см. рис. 25-37) или при слишком обширных повреждениях подверженная ошибкам репарация через повреждение. В последнем механизме задействована другая ДНК-полимераза (ДНК-полимераза V, кодируемая генами umuС и umuD), которая может реплицировать, хотя и с ошибками, участки ДНК с различными типами повреждений. Такой механизм называется репарацией, «подверженной ошибкам», из-за частого возникновения мутаций.

При застопоривании бактериальной репликативной вилки возможны два пути репарации. В отсутствие второй цепи информация, необходимая для точной репарации, должна поступить от гомологичной хромосомы. Таким образом, система репарации использует гомологичную генетическую рекомбинацию. Рекомбинационная репарация ДНК подробно рассматривается в разд. 25.3. При некоторых обстоятельствах реализуется второй путь репарации — подверженный ошибкам синтез ДНК через повреждение(TLS — от англ. translesion synthesis). При активации этого пути репарация ДНК становится существенно менее точной и связана с высокой частотой мутаций. У бактерий подверженный ошибкам синтез ДНК через повреждение — часть стрессового ответа клетки на сильное повреждение ДНК, который называют SOS-ответом. Некоторые SOS-белки, например, уже описанные UvrA и UvrB (табл. 25-6), всегда присутствуют в клетке, но при запуске SOS-ответа их уровень значительно повышается. В подверженной ошибкам репарации участвуют и другие SOS-белки, в том числе UmuC и UmuD («Umu» — от англ. immutable — нему тируемый; без функционального гена ити подверженная ошибкам репарация отсутствует). В SOS-регулируемом процессе белок UmuD превращается в более короткий фрагмент UmuD’, который в комплексе с UmuC образует специализированную ДНК-полимеразу (ДНК-полимеразу V), способную восстанавливать многие повреждения ДНК, блокирующие репликацию. В дефектном участке правильное спаривание оснований часто оказывается невозможным, поэтому такая репликация через повреждение сопровождается ошибками.
Таблица 25-6. Гены, индуцируемые как часть SOS-ответа у E. Coli
|
Название гена Гены с известной функцией polB (dinA) |
Кодируемый белок и/или его роль в репарации ДНК Кодирует субъединицу ДНК-полимеразы III с полимеразной активностью, необходимую для возобновления рекомбинационной репарации |
|
uvrA uvrB |
Кодируют субъединицы UvrA и UvrB АВС-экзонуклеазы |
|
umuС umuD |
Кодируют ДНК-полимеразу V |
|
sulA |
Кодирует белок, ингибирующий клеточное деление, возможно, высвобождая время для репарации ДНК |
|
recA |
Кодирует RecA белок, необходимый для подверженной ошибка» репарации и рекомбинационной репарации |
|
dinB |
Кодирует ДНК-полимеразу IV |
|
himA |
Кодирует субъединицу клеточного фактора интеграции (IHF), участвующего в сайт-специфической рекомбинации, репликации, транспозиции фагов, регуляции экспрессии генов |
|
Гены метаболизма ДНК с неизвестной функцией в репарации |
|
|
ssb |
Кодирует белок, связывающийся с одноцепочечной ДНК (SSB) |
|
uvrD |
Кодирует ДНК-хеликазу II (раскручивает ДНК) |
|
recN |
Необходим для рекомбинационной репарации |
|
Гены, функция которых не установлена |
|
|
dinD |
|
|
dinF |
Некоторые из этих генов и их функции обсуждаются в гл. 28.
В этой главе мы многократно подчеркивали особое значение сохранности генома, и поэтому существование системы, повышающей вероятность возникновения мутаций, может показаться странным. Использование этой системы можно воспринимать, как некий шаг отчаянья. Гены umuС и umuD полностью индуцируются только на поздней стадии SOS-ответа, они не активируются для синтеза, инициируемого расщеплением UmuD, до тех пор, пока уровень повреждений ДНК не становится слишком высоким, и блокируются все репликативные вилки. Мутации, возникающие в результате такой репликации, приводят к гибели одних клеток и опасны для других, но эту биологическую цену организм платит за то, чтобы не допустить прекращения репликации, поскольку это позволяет выжить хотя бы нескольким мутантным дочерним клеткам.
Наряду с ДНК-полимеразы V для репликации через поврежденный участок необходим белок RecA. Филаменты RecA, связанные с одноцепочечной ДНК в одном участке хромосомы, могут активировать комплексы ДНК-полимеразы V, связанные в отдаленных участках хромосомы. Реализации этого так называемого транс-механизма способствует образование петель, сближающих в пространстве удаленные друг от друга последовательности хромосом. В ходе SOS-ответа индуцируется еще одна ДНК-полимераза — ДНК-полимераза IV (продукт гена dinB). Репликация под действием ДНК-полимеразы IV также происходит с большим количеством ошибок. Бактериальные ДНК-полимеразы IV и V относятся к семейству TLS-полимераз, найденному у всех организмов. Эти ферменты не имеют корректирующей экзонуклеазной активности, а точность в выборе оснований при репликации в 102 раз ниже, что уменьшает точность репликации в целом примерно до одной ошибки на 1000 нуклеотидов.
У млекопитающих есть много ДНК-полимераз из семейства TLS-полимераз, отличающихся низкой точностью синтеза. Однако присутствие этих ферментов не является причиной повышенной частоты мутаций, поскольку большинство из них выполняют особые функции при репарации. ДНК-полимераза г), например, относится к TLS-полимеразам и обнаружена у всех эукариот. Она осуществляет синтез через поврежденные участки, главным образом, через циклобутановые Т-Т-димеры. В этом случае образуется немного мутаций, поскольку фермент предпочтительно вставляет два остатка А напротив связанных остатков Т. У эукариот несколько полимераз, включая ДНК-полимеразы β, յ и λ, выполняют специальные функции с низкой точностью при эксцизионной репарации оснований. Каждый из этих ферментов наряду с полимеразной активностью имеет активность 5′-дезоксирибозофосфат-лиазы. После удаления основания гликозилазой и расщепления остова молекулы АР-эндонуклеазой эти ферменты удаляют АР-сайт (5′-дезоксирибозофосфат) и заполняют очень короткую брешь. Количество мутаций, возникающих при действии ДНК-полимеразы n, невелико по той причине, что она синтезирует очень короткие участки ДНК (часто в один нуклеотид).
В результате исследования систем репарации клеточной ДНК стало понятно, что метаболизм ДНК поддерживает целостность генома с помощью многочисленных и часто избыточных систем. В геноме человека более 130 генов кодируют белки, связанные с репарацией ДНК. Во многих случаях потеря функции одного гена приводит к нестабильности генома и повышению вероятности опухолевых заболеваний (доп. 25-1). Эти системы репарации часто работают в комплексе с системами репликации ДНК и дополняются системами рекомбинации, которые будут рассмотрены ниже.
Дополнение 25-1. МЕДИЦИНА. Репарация ДНК и рак
Рак у человека возникает, когда определенные гены, регулирующие нормальное клеточное деление (онкогены и опухолевые супрессоры; см. гл. 12 в т. 1), перестают нормально функционировать, изменяются или активируются не вовремя. В результате рост клеток может выйти из-под контроля — возникает опухоль. Гены, контролирующие клеточное деление, могут быть повреждены спонтанной мутацией или перекрываться инвазией опухолевого вируса (гл. 26). Неудивительно, что изменения в генах репарации ДНК, приводящие к усилению мутагенеза, могут значительно повысить индивидуальную чувствительность к раку. Все дефекты генов белков, участвующих в эксцизионной репарации оснований, репарации ошибочно спаренных оснований, рекомбинационной репарации и SOS-репарации, связаны с онкологическими заболеваниями. Итак, от репарации ДНК может зависеть жизнь и смерть.
Эксцизионная репарация оснований у человека, в отличие от бактерий, нуждается в большем числе белков, хотя в целом механизмы очень похожи. Генетические дефекты, сопровождающиеся инактивацией экс- цизионной репарации оснований, связаны с несколькими генетическими заболеваниями, среди которых лучше других исследована пигментная ксеродерма, или ХР (от лат. xeroderma pigmentosum). Поскольку у человека эксцизионная репарация оснований — единственный способ репарации пиримидиновых димеров, люди с ХР очень чувствительны к свету и часто заболевают раком кожи, который индуцируется солнечным светом. У большинства больных ХР также выражены неврологические расстройства, предположительно из-за отсутствия у них репарации определенных повреждений, вызванных высокой активностью окислительного метаболизма в нейронах. Дефекты в генах, кодирующих один из семи (как минимум) разных белковых компонентов системы эксцизионной репарации оснований, могут привести к семи генетическим вариантам ХР (от ХРА до XPG). Некоторые из этих белков (особенно те, которые повреждены при ХРВ, XPD и XPG) также участвуют в сопряженной с транскрипцией репарации окислительных повреждений, как описано в гл. 26.
У большинства микроорганизмов есть дополнительные пути репарации циклобутановых пиримидиновых димеров — они могут использовать ДНК- фотолиазную активность, а иногда и эксцизионную репарацию оснований в качестве альтернативного механизма эксцизионной репарации нуклеотидов, но у человека и других плацентарных млекопитающих такой возможности нет. Отсутствие альтернатив эксцизионной репарации нуклеотидов для удаления пиримидиновых димеров вызвало предположение, что на ранней стадии эволюции млекопитающие были мелкими животными, были покрыты мехом и вели ночной образ жизни, не очень нуждаясь в репарации повреждений, вызванных УФ-облучением. Однако у млекопитающих есть путь синтеза, позволяющий обойти циклобутановые пиримидиновые димеры, и в нем участвует ДНК-полимераза n. Этот фермент предпочтительно вставляет два остатка А напротив пиримидинового димера Т-Т, минимизируя вероятность мутации. Люди, у которых в силу генетических нарушений не работает ДНК-полимераза n, страдают заболеванием, похожим на пигментную ксеродерму (ХР-вариант V, или XPV). Клинические проявления XPV напоминают симптомы классической формы ХР, хотя уровень мутаций при XPV бывает выше, когда клетки подвергаются воздействию УФ-облучения. По-видимому, в нормальных клетках система эксцизионной репарации нуклеотидов работает совместно с ДНК-полимеразой n, восстанавливая и или обходя пиримидиновые димеры, что необходимо для нормального клеточного роста и репликации ДНК. УФ-облучение приводит к появлению множества пиримидиновых димеров, и для поддержания репликации хотя бы часть из них приходится обходить путем синтеза через повреждение. Если одна из систем отсутствует, ее частично компенсирует другая система. Отсутствие активности полимеразы n приводит к остановке репликативных вилок и исправлению повреждений от УФ-света TLS-полимеразами, которые допускают гораздо больше ошибок. И если отсутствуют другие системы репарации ДНК, увеличение числа мутаций часто приводит к раку.
Один из распространенных наследуемых синдромов предрасположенности к возникновению злокачественных опухолей — наследственный неполипозный рак толстой кишки (ННРТК). Этот синдром связан с нарушением репарации ошибочно спаренных оснований. В клетках человека и других эукариот есть несколько белковых аналогов бактериальных белков MutL и MutS (см. рис. 25-23). Вероятность развития ННРТК могут повысить дефекты по крайней мерс в пяти разных генах репарации ошибочно спаренных оснований. Особенно распространены дефекты в hMLH1 (человеческий гомолог 1 MutL) и hMSH2 (человеческий гомолог 2 MutS). У человека с ННРТК рак обычно развивается в раннем возрасте, и чаще всего это бывает рак толстой кишки.
Большинство случаев рака молочной железы у женщин возникает без известной генетической предрасположенности. Однако в 10% случаев это заболевание связано с наследственным дефектом двух генов — BRCA1 и BRCA2. У человека белки BRCA1 и BRCA2 — это очень большие молекулы (1834 и 3418 аминокислотных остатков соответственно), которые взаимодействуют с многими другими белками, участвующими в транскрипции, поддержании хромосом, репарации ДНК и контроле клеточного цикла. Кроме того, белок BRCA2 связан с рекомбинационной репарацией двухцепочечных разрывов. Однако точная молекулярная функция BRCA1 и BRCA2 в этих клеточных процессах до сих пор не установлена. Вероятность заболевания раком груди у женщин с дефектами в гене BRCA1 или BRCA2 превышает 80%.
Краткое содержание раздела 25.2 Репарация ДНК
■ Клетки имеют несколько систем репарации ДНК. У Е. coli репарация ошибочно спаренных оснований осуществляется при временном отсутствии метилирования последовательностей (5′) GATС во вновь синтезированной цепи.
■ Системы эксцизионной репарации распознают и устраняют повреждения, вызванные факторами внешней среды (такими, как радиация и алкилирующие агенты) и спонтанными реакциями нуклеотидов. Некоторые системы репарации распознают и вырезают только поврежденные или неправильные основания, оставляя АР-сайт (лишенный основания сайт) в ДНК, который вырезается и заполняется новой ДНК под действием других ферментов.
■ Системы эксцизионной репарации нуклеотидов распознают и удаляют различные крупные повреждения и пиримидиновые димеры. Они вырезают участок цепи ДНК, содержащий повреждение, и оставляют брешь, которую заделывают ДНК-полимераза и лигаза.
■ Некоторые повреждения ДНК ликвидируются в реакциях, обратных тем, которые вызывали дефект: пиримидиновые димеры превращаются в мономерные пиримидины с помощью фотолиазы, а метальная группа О6— метилгуанина удаляется метилтрансферазой.
■ При очень сильных повреждениях бактериальной ДНК происходит синтез новой последовательности через повреждение с участием TLS-полимеразы, допускающей достаточно много ошибок. Эукариоты имеют похожие полимеразы, выполняющие специализированные функции в репарации ДНК, что минимизирует количество мутаций.